

Nutrient-driven O-GlcNAc in proteostasis and neurodegeneration
- PMID: 29049853
- PMCID: PMC5735008
- DOI: 10.1111/jnc.14242
Nutrient-driven O-GlcNAc in proteostasis and neurodegeneration
Abstract
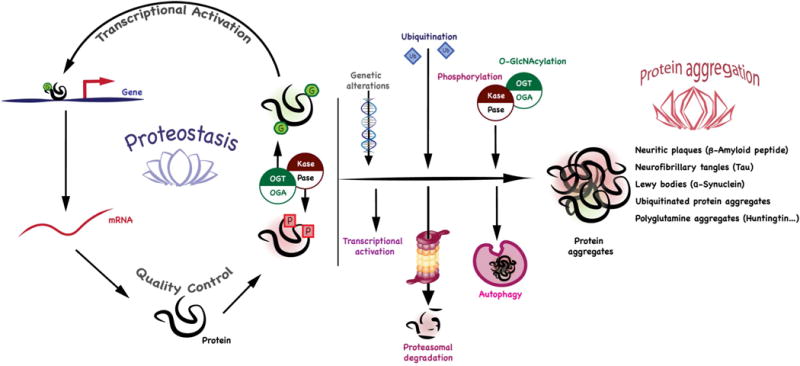
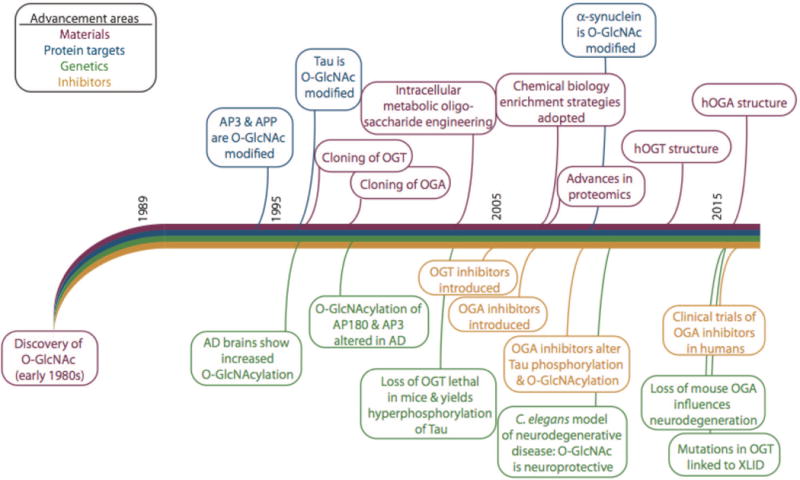
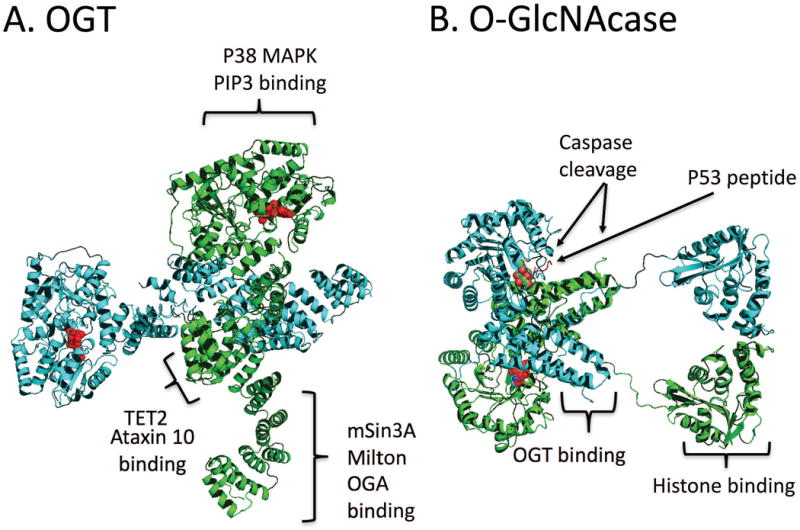
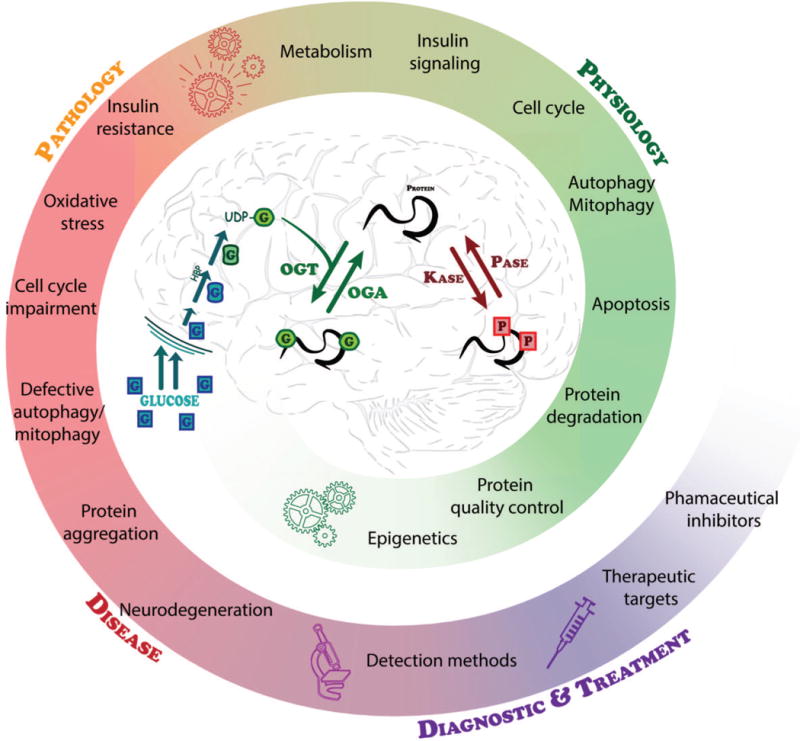
Proteostasis is essential in the mammalian brain where post-mitotic cells must function for decades to maintain synaptic contacts and memory. The brain is dependent on glucose and other metabolites for proper function and is spared from metabolic deficits even during starvation. In this review, we outline how the nutrient-sensitive nucleocytoplasmic post-translational modification O-linked N-acetylglucosamine (O-GlcNAc) regulates protein homeostasis. The O-GlcNAc modification is highly abundant in the mammalian brain and has been linked to proteopathies, including neurodegenerative diseases such as Alzheimer's, Parkinson's, and Huntington's. C. elegans, Drosophila, and mouse models harboring O-GlcNAc transferase- and O-GlcNAcase-knockout alleles have helped define the role O-GlcNAc plays in development as well as age-associated neurodegenerative disease. These enzymes add and remove the single monosaccharide from protein serine and threonine residues, respectively. Blocking O-GlcNAc cycling is detrimental to mammalian brain development and interferes with neurogenesis, neural migration, and proteostasis. Findings in C. elegans and Drosophila model systems indicate that the dynamic turnover of O-GlcNAc is critical for maintaining levels of key transcriptional regulators responsible for neurodevelopment cell fate decisions. In addition, pathways of autophagy and proteasomal degradation depend on a transcriptional network that is also reliant on O-GlcNAc cycling. Like the quality control system in the endoplasmic reticulum which uses a 'mannose timer' to monitor protein folding, we propose that cytoplasmic proteostasis relies on an 'O-GlcNAc timer' to help regulate the lifetime and fate of nuclear and cytoplasmic proteins. O-GlcNAc-dependent developmental alterations impact metabolism and growth of the developing mouse embryo and persist into adulthood. Brain-selective knockout mouse models will be an important tool for understanding the role of O-GlcNAc in the physiology of the brain and its susceptibility to neurodegenerative injury.
Keywords: Alzheimer's; Glucose; Neurodegeneration; O-GlcNAc; O-linked N-acetylglucosamine; Therapeutics.
Published 2017. This article is a U.S. Government work and is in the public domain in the USA.
Conflict of interest statement
The authors have no conflicts to declare
Figures
References
-
- Akimoto Y, Comer FI, Cole RN, Kudo A, Kawakami H, Hirano H, Hart GW. Localization of the O-GlcNAc transferase and O-GlcNAc-modified proteins in rat cerebellar cortex. Brain Res. 2003;966:194–205. - PubMed
-
- Akimoto Y, Kreppel LK, Hirano H, Hart GW. Localization of the O-linked N-acetylglucosamine transferase in rat pancreas. Diabetes. 1999;48:2407–2413. - PubMed
-
- Akimoto Y, Kreppel LK, Hirano H, Hart GW. Increased O-GlcNAc transferase in pancreas of rats with streptozotocin-induced diabetes. Diabetologia. 2000;43:1239–1247. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
