

The dual role of poly(ADP-ribose) polymerase-1 in modulating parthanatos and autophagy under oxidative stress in rat cochlear marginal cells of the stria vascularis
- PMID: 29049980
- PMCID: PMC5641921
- DOI: 10.1016/j.redox.2017.10.002
The dual role of poly(ADP-ribose) polymerase-1 in modulating parthanatos and autophagy under oxidative stress in rat cochlear marginal cells of the stria vascularis
Abstract
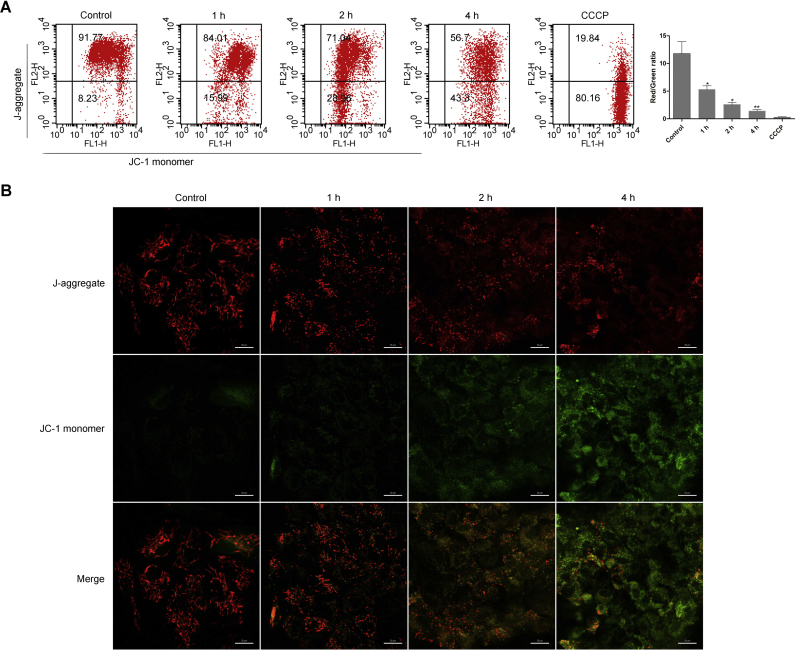
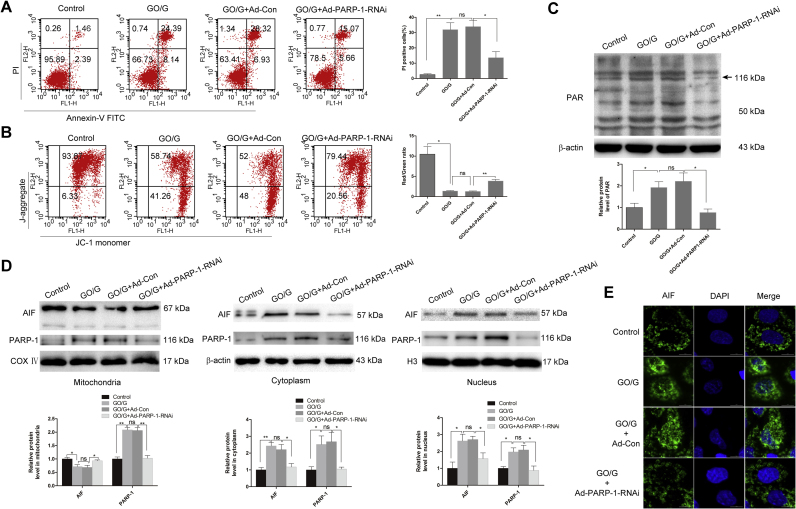
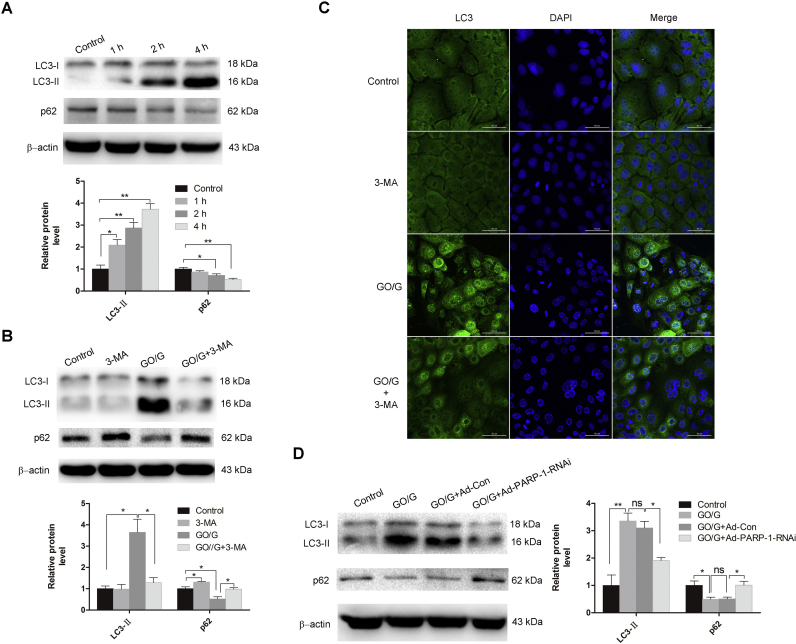
Oxidative stress is reported to regulate several apoptotic and necrotic cell death pathways in auditory tissues. Poly(ADP-ribose) polymerase-1 (PARP-1) can be activated under oxidative stress, which is the hallmark of parthanatos. Autophagy, which serves either a pro-survival or pro-death function, can also be stimulated by oxidative stress, but the role of autophagy and its relationship with parthanatos underlying this activation in the inner ear remains unknown. In this study, we established an oxidative stress model in vitro by glucose oxidase/glucose (GO/G), which could continuously generate low concentrations of H2O2 to mimic continuous exposure to H2O2 in physiological conditions, for investigation of oxidative stress-induced cell death mechanisms and the regulatory role of PARP-1 in this process. We observed that GO/G induced stria marginal cells (MCs) death via upregulation of PARP-1 expression, accumulation of polyADP-ribose (PAR) polymers, decline of mitochondrial membrane potential (MMP) and nuclear translocation of apoptosis-inducing factor (AIF), which all are biochemical features of parthanatos. PARP-1 knockdown rescued GO/G-induced MCs death, as well as abrogated downstream molecular events of PARP-1 activation. In addition, we demonstrated that GO/G stimulated autophagy and PARP-1 knockdown suppressed GO/G-induced autophagy in MCs. Interestingly, autophagy suppression by 3-Methyladenine (3-MA) accelerated GO/G-induced parthanatos, indicating a pro-survival function of autophagy in GO/G-induced MCs death. Taken together, these data suggested that PARP-1 played dual roles by modulating parthanatos and autophagy in oxidative stress-induced MCs death, which may be considered as a promising therapeutic target for ameliorating oxidative stress-related hearing disorders.
Keywords: Autophagy; Glucose oxidase; Marginal cells; Oxidative stress; PARP-1; Parthanatos.
Copyright © 2017 The Authors. Published by Elsevier B.V. All rights reserved.
Figures




Similar articles
-
PARP1 inhibition prevents oxidative stress in age-related hearing loss via PAR-Ca2+-AIF axis in cochlear strial marginal cells.Free Radic Biol Med. 2024 Aug 1;220:222-235. doi: 10.1016/j.freeradbiomed.2024.05.020. Epub 2024 May 10. Free Radic Biol Med. 2024. PMID: 38735540
-
Arsenite-induced cytotoxicity is regulated by poly-ADP ribose polymerase 1 activation and parthanatos in p53-deficient H1299 cells: The roles of autophagy and p53.Biochem Biophys Res Commun. 2023 May 14;656:78-85. doi: 10.1016/j.bbrc.2023.03.018. Epub 2023 Mar 14. Biochem Biophys Res Commun. 2023. PMID: 36958258
-
LPS protects macrophages from AIF-independent parthanatos by downregulation of PARP1 expression, induction of SOD2 expression, and a metabolic shift to aerobic glycolysis.Free Radic Biol Med. 2019 Feb 1;131:184-196. doi: 10.1016/j.freeradbiomed.2018.11.034. Epub 2018 Nov 28. Free Radic Biol Med. 2019. PMID: 30502458
-
Poly (ADP-Ribose) polymerase 1 and parthanatos in neurological diseases: From pathogenesis to therapeutic opportunities.Neurobiol Dis. 2023 Oct 15;187:106314. doi: 10.1016/j.nbd.2023.106314. Epub 2023 Oct 1. Neurobiol Dis. 2023. PMID: 37783233 Review.
-
Poly(ADP-ribose) signals to mitochondrial AIF: a key event in parthanatos.Exp Neurol. 2009 Aug;218(2):193-202. doi: 10.1016/j.expneurol.2009.03.020. Epub 2009 Mar 28. Exp Neurol. 2009. PMID: 19332058 Free PMC article. Review.
Cited by
-
Rhinovirus and Cell Death.Viruses. 2021 Apr 7;13(4):629. doi: 10.3390/v13040629. Viruses. 2021. PMID: 33916958 Free PMC article. Review.
-
Flow Cytometry Reveals the Nature of Oncotic Cells.Int J Mol Sci. 2019 Sep 6;20(18):4379. doi: 10.3390/ijms20184379. Int J Mol Sci. 2019. PMID: 31489916 Free PMC article.
-
Inhibition of AKT induces p53/SIRT6/PARP1-dependent parthanatos to suppress tumor growth.Cell Commun Signal. 2022 Jun 17;20(1):93. doi: 10.1186/s12964-022-00897-1. Cell Commun Signal. 2022. PMID: 35715817 Free PMC article.
-
Cellular autophagy, the compelling roles in hearing function and dysfunction.Front Cell Neurosci. 2022 Sep 30;16:966202. doi: 10.3389/fncel.2022.966202. eCollection 2022. Front Cell Neurosci. 2022. PMID: 36246522 Free PMC article. Review.
-
Cochlear Marginal Cell Pyroptosis Is Induced by Cisplatin via NLRP3 Inflammasome Activation.Front Immunol. 2022 Apr 20;13:823439. doi: 10.3389/fimmu.2022.823439. eCollection 2022. Front Immunol. 2022. PMID: 35529876 Free PMC article.
References
-
- Schuknecht H.F., Gacek M.R. Cochlear pathology in presbycusis. Ann. Otol. Rhinol. Laryngol. 1993;102:1–16. - PubMed
-
- Tavanai E., Mohammadkhani G. Role of antioxidants in prevention of age-related hearing loss: a review of literature. Eur. Arch. Otorhinolaryngol. 2017;274:1821–1834. - PubMed
-
- Rigoulet M., Yoboue E.D., Devin A. Mitochondrial ROS generation and its regulation: mechanisms involved in H(2)O(2) signaling. Antioxid. Redox Signal. 2011;14:459–468. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
