

Cyclic AMP-Responsive Element-Binding Protein (CREB) is Critical in Autoimmunity by Promoting Th17 but Inhibiting Treg Cell Differentiation
- PMID: 29050947
- PMCID: PMC5704088
- DOI: 10.1016/j.ebiom.2017.10.010
Cyclic AMP-Responsive Element-Binding Protein (CREB) is Critical in Autoimmunity by Promoting Th17 but Inhibiting Treg Cell Differentiation
Erratum in
-
Corrigendum to 'Cyclic AMP-Responsive element-binding protein (CREB) is critical in autoimmunity by promoting Th17 but inhibiting treg cell differentiation': [EBioMedicine 25 (2017) 165-174].EBioMedicine. 2020 Jul;57:102874. doi: 10.1016/j.ebiom.2020.102874. Epub 2020 Jul 7. EBioMedicine. 2020. PMID: 32650272 Free PMC article. No abstract available.
Abstract
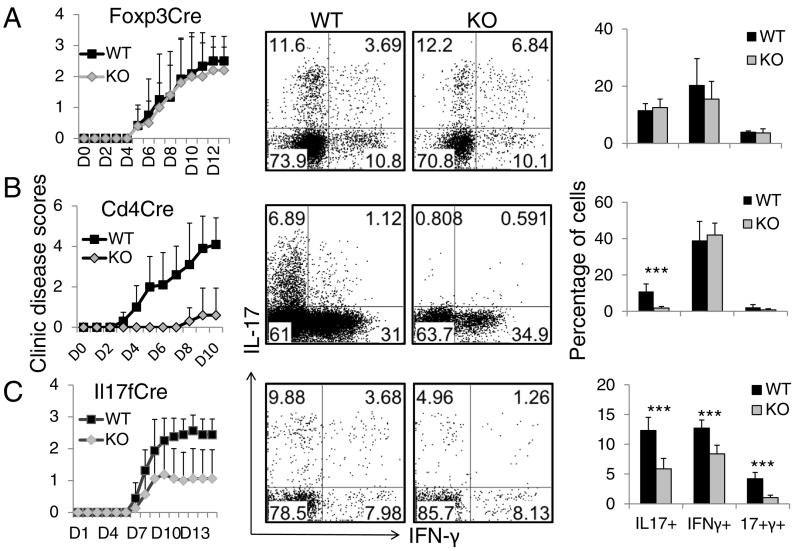
The molecular mechanisms that govern differential T cell development into pro-inflammatory Th17 vs. regulatory T (Treg) cells remain unclear. Here, we show that selective deletion of CREB in T cells or Th17 cells impaired Th17 cell differentiation in vitro and in vivo, and led to resistance to autoimmune diseases. Mechanistically, CREB, activated by CD3-PKC-ϴ signaling, plays a key role in regulating Th17 cell differentiation, at least in part through directly binding to the Il17-Il17f gene locus. Unexpectedly, although dispensable for FOXP3 expression and for the homeostasis and suppressive function of thymus-derived Treg cells, CREB negatively regulates the survival of TGF-β-induced Treg cells, and deletion of CREB resulted in increased FOXP3+ Treg cells in the intestine and protection in a colitis model. Thus, CREB is critical in autoimmune diseases by promoting Th17 cell and inhibiting de novo Treg cell generation.
Keywords: Autoimmune diseases; CREB; Th17 cells; Treg cells.
Copyright © 2017 The Author(s). Published by Elsevier B.V. All rights reserved.
Figures






Comment in
-
Dual Mechanisms for Balancing Th17 and Treg Cell Fate by CREB.EBioMedicine. 2017 Nov;25:20-21. doi: 10.1016/j.ebiom.2017.10.031. Epub 2017 Nov 10. EBioMedicine. 2017. PMID: 29129697 Free PMC article.
References
-
- Altman A., Kong K.F. PKCtheta: a new target for selective immunosuppression. Expert. Rev. Clin. Immunol. 2012;8:205–208. - PubMed
-
- Barton K., Muthusamy N., Chanyangam M., Fischer C., Clendenin C., Leiden J.M. Defective thymocyte proliferation and IL-2 production in transgenic mice expressing a dominant-negative form of CREB. Nature. 1996;379:81–85. - PubMed
-
- Boniface K., Bak-Jensen K.S., Li Y., Blumenschein W.M., McGeachy M.J., McClanahan T.K., McKenzie B.S., Kastelein R.A., Cua D.J., de Waal Malefyt R. Prostaglandin E2 regulates Th17 cell differentiation and function through cyclic AMP and EP2/EP4 receptor signaling. J. Exp. Med. 2009;206:535–548. - PMC - PubMed
-
- Dodon M.D., Li Z., Hamaia S., Gazzolo L. Tax protein of human T-cell leukaemia virus type 1 induces interleukin 17 gene expression in T cells. J. Gen. Virol. 2004;85:1921–1932. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
