

HMGA1 is a novel transcriptional regulator of the FoxO1 gene
- PMID: 29052178
- PMCID: PMC5845622
- DOI: 10.1007/s12020-017-1445-8
HMGA1 is a novel transcriptional regulator of the FoxO1 gene
Abstract
Purpose: The forkhead transcription factor (FoxO1) is a master transcriptional regulator of fundamental cellular processes ranging from cell proliferation and differentiation to inflammation and metabolism. However, despite its relevance, the mechanism(s) underlying FoxO1 gene regulation are largely unknown. We have previously shown that the chromatin factor high-mobility group A1 (HMGA1) plays a key role in the transcriptional regulation of glucose-responsive genes, including some that are involved in FoxO1-mediated glucose metabolism. Here we investigated the impact of HMGA1 on FoxO1 gene expression.
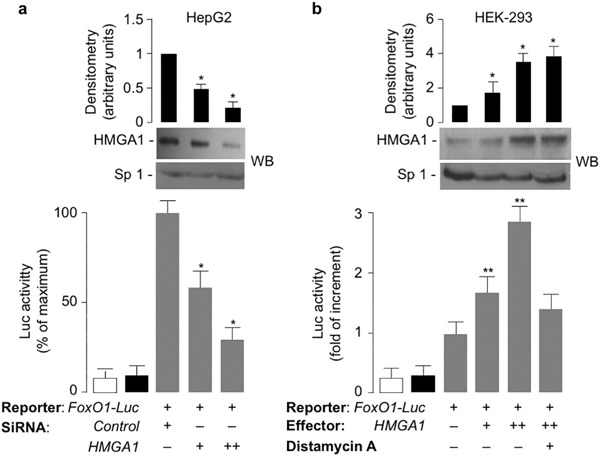
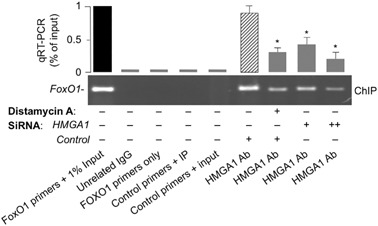
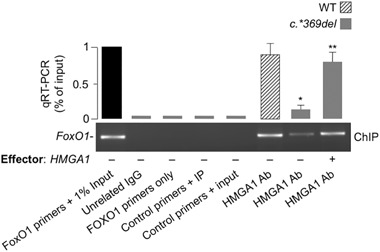
Methods: FoxO1 protein and gene expression studies were performed by Western blot analysis combined with qRT-PCR of material from human cultured cells and EBV-transformed lymphoblasts, and from primary cultured hepatocytes from wild-type and Hmga1 -/- mice. Reporter gene assays and chromatin immunoprecipitation for binding of HMGA1 to the endogenous FoxoO1 locus were performed in cells overexpressing HMGA1 and in cells pretreated with siRNA targeting HMGA1.
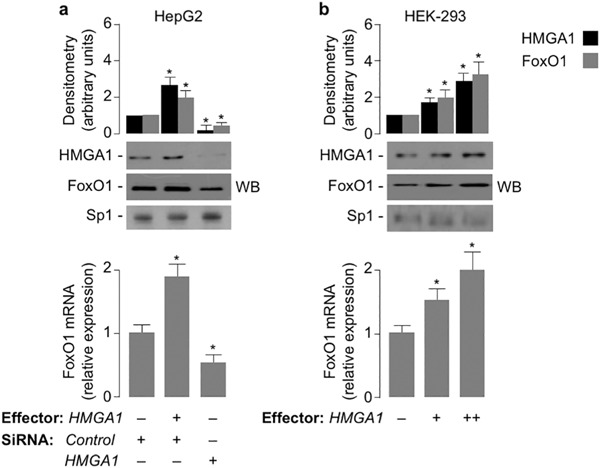
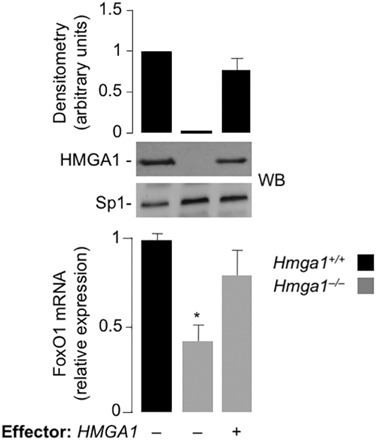
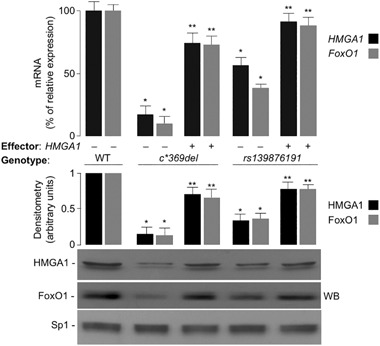
Results: HMGA1 increased FoxO1 mRNA and protein expression in vitro, in cultured HepG2 and HEK-293 cells by binding FoxO1 gene promoter, thereby activating FoxO1 gene transcription. Forced expression of HMGA1 in primary cultured hepatocytes from Hmga1 -/- mice and in EBV-transformed lymphoblasts from subjects with reduced expression of endogenous HMGA1 increased FoxO1 mRNA and protein levels.
Conclusion: These findings may contribute to the understanding of FoxO1 gene regulation and its role in metabolism.
Keywords: DNA/chromatin interaction; FoxO1; Gene transcription; HMGA1; Insulin signaling.
Conflict of interest statement
Conflict of interest
The authors declare that they have no competing interests.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Informed consent
For this type of study formal consent is not required.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
