

Social behaviour shapes hypothalamic neural ensemble representations of conspecific sex
- PMID: 29052632
- PMCID: PMC5674977
- DOI: 10.1038/nature23885
Social behaviour shapes hypothalamic neural ensemble representations of conspecific sex
Abstract
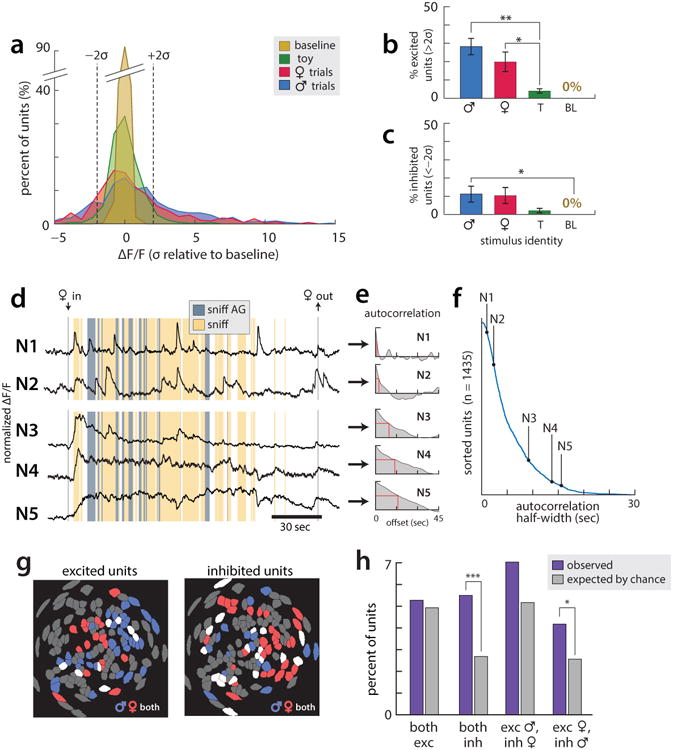
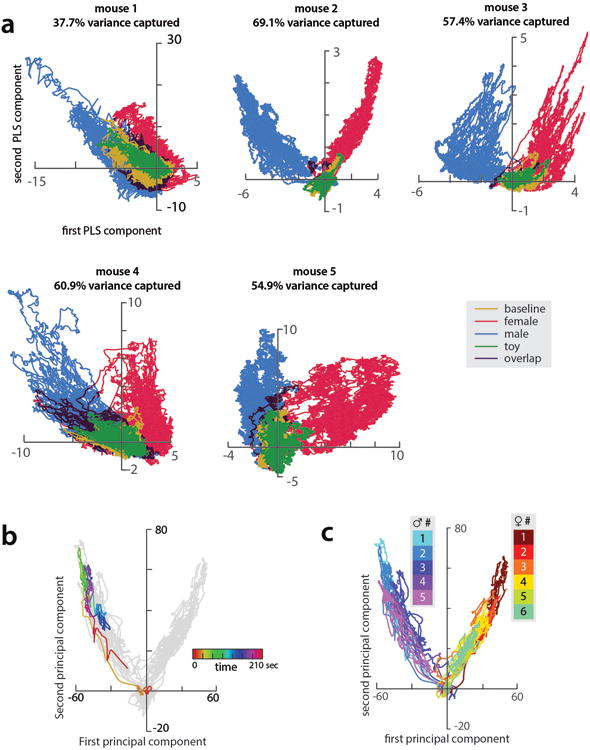
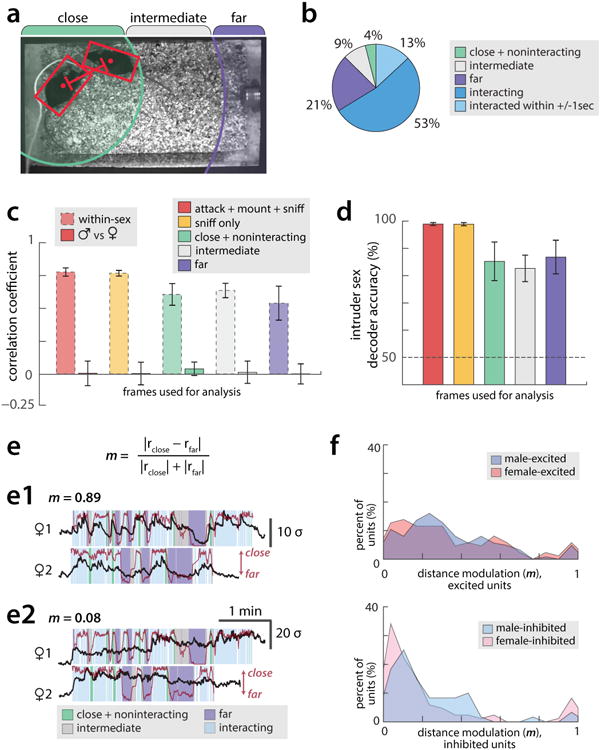
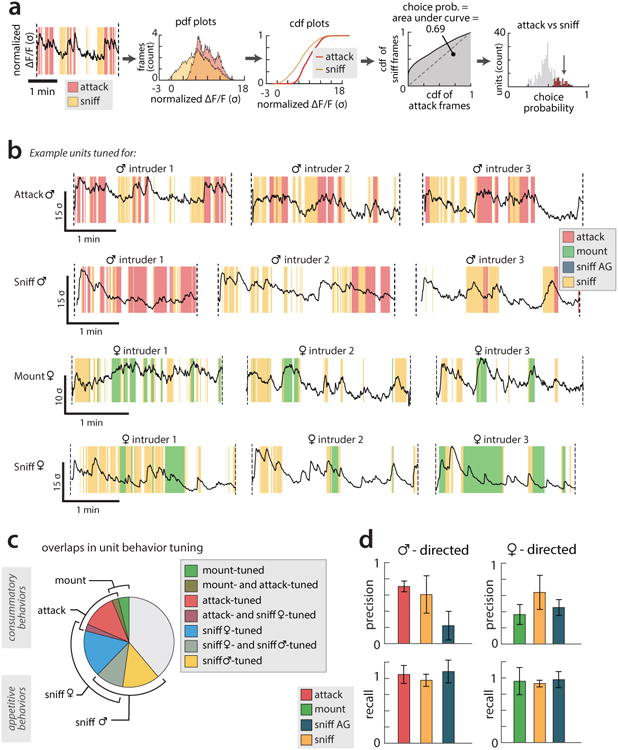
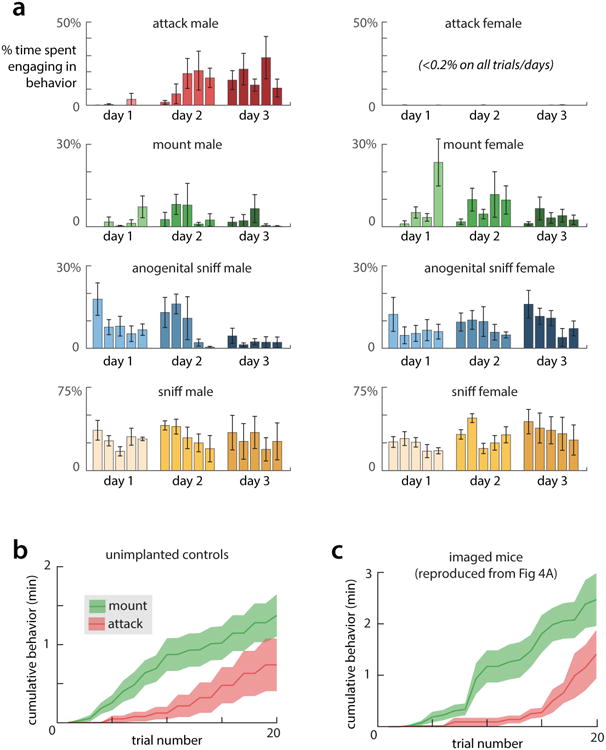
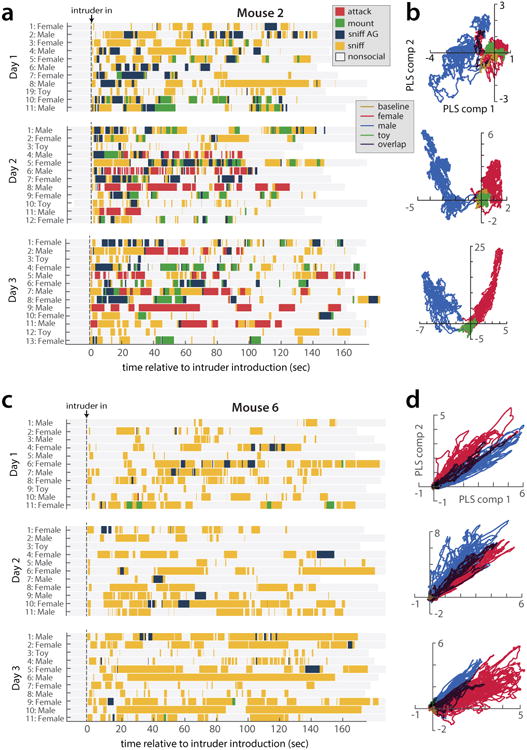
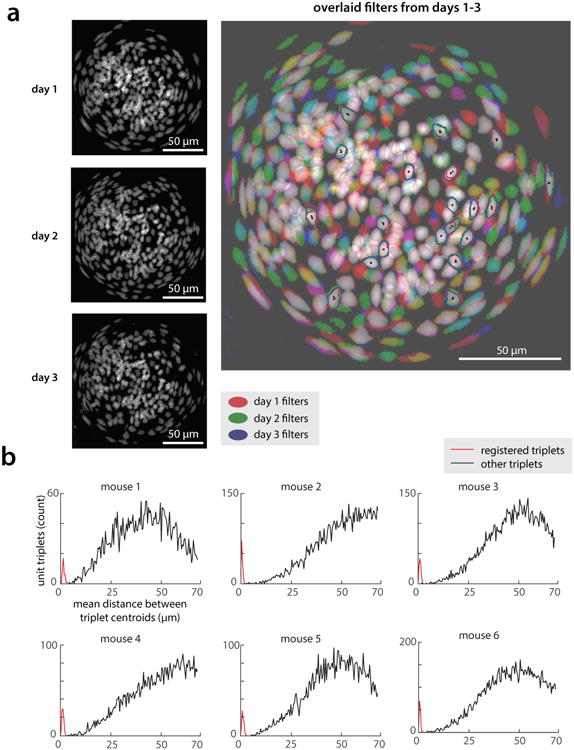
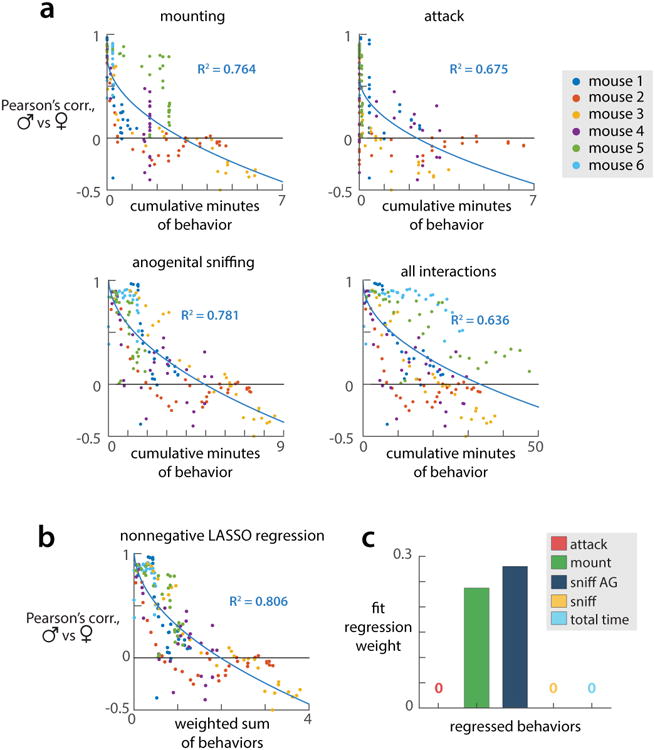
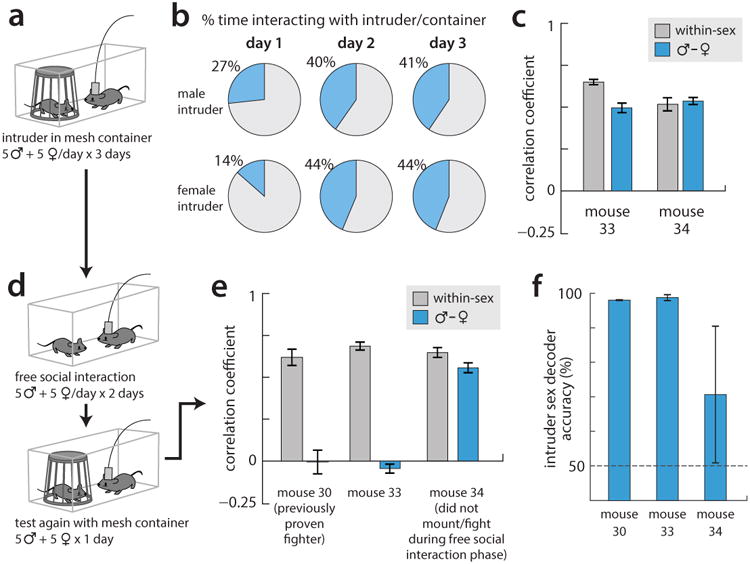
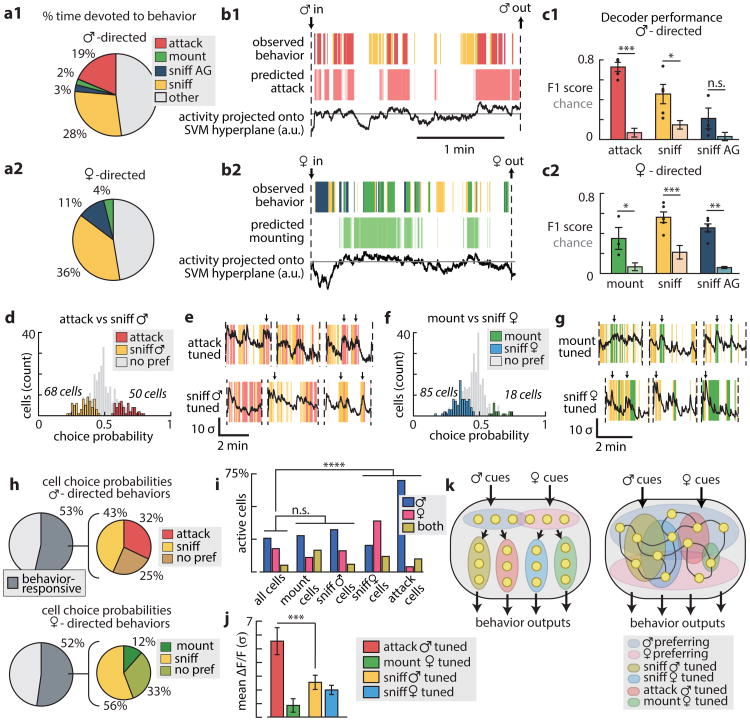
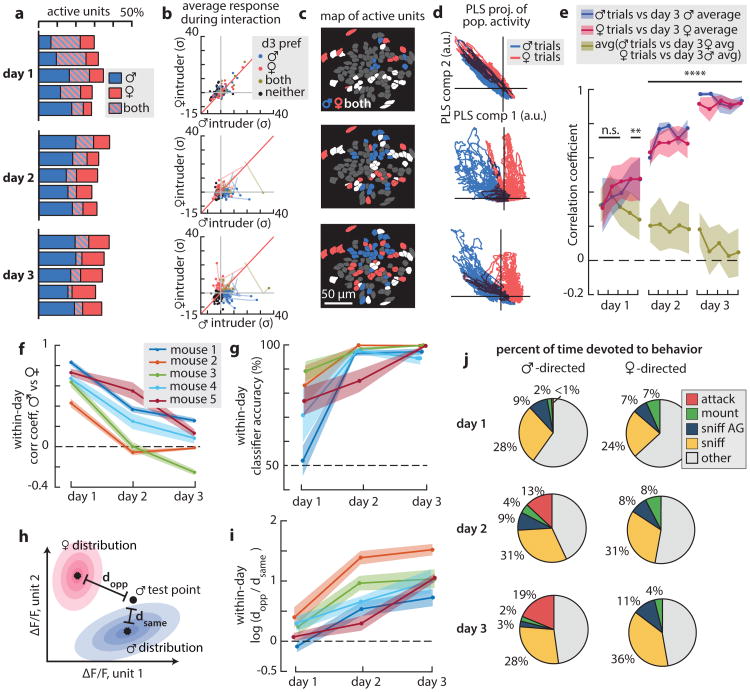
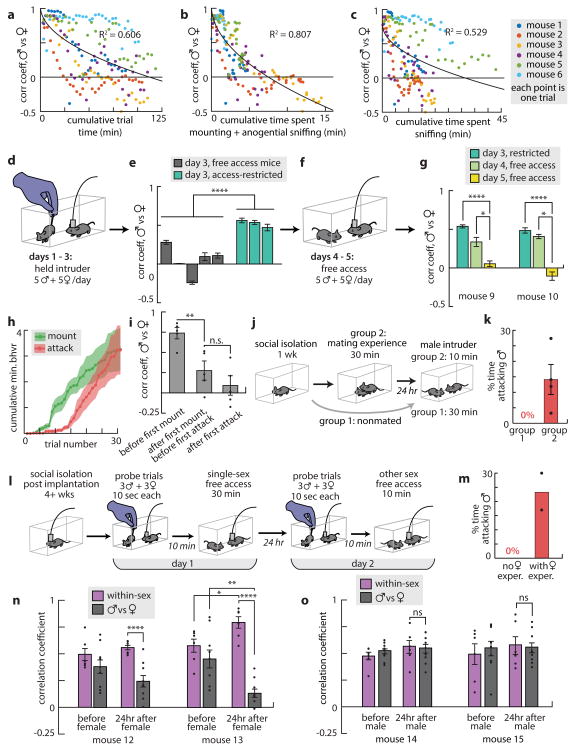
All animals possess a repertoire of innate (or instinctive) behaviours, which can be performed without training. Whether such behaviours are mediated by anatomically distinct and/or genetically specified neural pathways remains unknown. Here we report that neural representations within the mouse hypothalamus, that underlie innate social behaviours, are shaped by social experience. Oestrogen receptor 1-expressing (Esr1+) neurons in the ventrolateral subdivision of the ventromedial hypothalamus (VMHvl) control mating and fighting in rodents. We used microendoscopy to image Esr1+ neuronal activity in the VMHvl of male mice engaged in these social behaviours. In sexually and socially experienced adult males, divergent and characteristic neural ensembles represented male versus female conspecifics. However, in inexperienced adult males, male and female intruders activated overlapping neuronal populations. Sex-specific neuronal ensembles gradually separated as the mice acquired social and sexual experience. In mice permitted to investigate but not to mount or attack conspecifics, ensemble divergence did not occur. However, 30 minutes of sexual experience with a female was sufficient to promote the separation of male and female ensembles and to induce an attack response 24 h later. These observations uncover an unexpected social experience-dependent component to the formation of hypothalamic neural assemblies controlling innate social behaviours. More generally, they reveal plasticity and dynamic coding in an evolutionarily ancient deep subcortical structure that is traditionally viewed as a 'hard-wired' system.
Figures


References
-
- Tinbergen N. The study of instinct. Clarendon Press; 1951.
-
- Lorenz K. On Aggression. Harcourt, Brace & World; 1966.
-
- Newman SW. The medial extended amygdala in male reproductive behavior. A node in the mammalian social behavior network. Annals of the New York Academy of Sciences. 1999;877:242–257. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
