

Selective base excision repair of DNA damage by the non-base-flipping DNA glycosylase AlkC
- PMID: 29054852
- PMCID: PMC5753038
- DOI: 10.15252/embj.201797833
Selective base excision repair of DNA damage by the non-base-flipping DNA glycosylase AlkC
Abstract
DNA glycosylases preserve genome integrity and define the specificity of the base excision repair pathway for discreet, detrimental modifications, and thus, the mechanisms by which glycosylases locate DNA damage are of particular interest. Bacterial AlkC and AlkD are specific for cationic alkylated nucleobases and have a distinctive HEAT-like repeat (HLR) fold. AlkD uses a unique non-base-flipping mechanism that enables excision of bulky lesions more commonly associated with nucleotide excision repair. In contrast, AlkC has a much narrower specificity for small lesions, principally N3-methyladenine (3mA). Here, we describe how AlkC selects for and excises 3mA using a non-base-flipping strategy distinct from that of AlkD. A crystal structure resembling a catalytic intermediate complex shows how AlkC uses unique HLR and immunoglobulin-like domains to induce a sharp kink in the DNA, exposing the damaged nucleobase to active site residues that project into the DNA This active site can accommodate and excise N3-methylcytosine (3mC) and N1-methyladenine (1mA), which are also repaired by AlkB-catalyzed oxidative demethylation, providing a potential alternative mechanism for repair of these lesions in bacteria.
Keywords: 3‐methyladenine; 3‐methylcytosine; DNA glycosylase; DNA repair; base excision repair.
© 2017 The Authors.
Figures
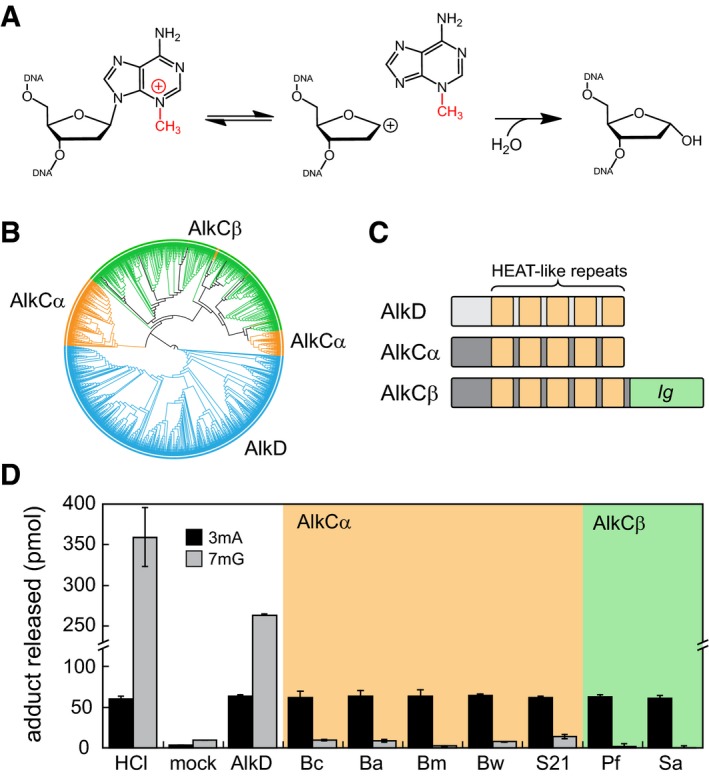
3mA excision reaction catalyzed by AlkC.
Phylogenetic tree of 779 AlkC (orange, green) and 764 AlkD (blue) protein sequences visualized using the iTOL web server (Letunic & Bork, 2011).
Schematic of AlkC and AlkD protein domains.
Release of 3mA (black bars) and 7mG (gray bars) from methylated genomic DNA after a 1‐h incubation with either HCl, no enzyme (mock), Bacillus cereus AlkD, or one of seven AlkC orthologs. Bc, Bacillus cereus; Ba, Bacillus anthracis; Bm, Bacillus mycoides; Bw, Bacillus weihenstephanensis; S21, Sphingobacterium sp. 21; Pf, Pseudomonas fluorescens; Sa, Streptomyces albus. Values are the mean ± SD (n = 3 for each). HCl and no‐enzyme controls provide upper and lower limits of 3mA and 7mG detection.
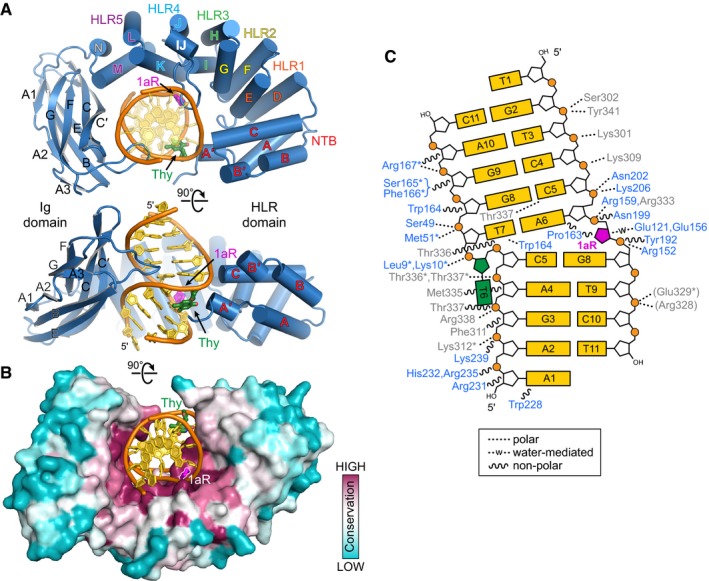
Two orthogonal views of the PfAlkC/1aR‐DNA complex crystal structure. The protein is colored blue, DNA gold, 1′‐aza‐2′,4′‐dideoxyribose (1aR) magenta, and opposite thymine green. NTB, N‐terminal helical bundle; HLR, HEAT‐like repeat.
AlkC sequence conservation (purple, high; cyan, low) superposed onto the protein surface.
Schematic illustration of AlkC‐DNA interactions. Dashed and wavy lines denote polar and non‐polar interactions, respectively. Residues from HLR and Ig‐like domains are blue and gray, respectively. Contacts to the protein backbone are marked with an asterisk, and symmetry‐related contacts are in parentheses.
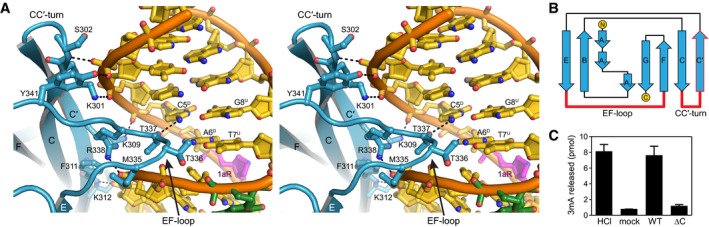
Stereoimage of the Ig‐like domain (blue) interactions with the major grove of DNA (gold). DNA‐interacting side chains are shown as sticks, and hydrogen bonds are shown as dashed lines. Superscripts in nucleotide labels refer to damaged (D) and undamaged (U) strands.
Topology of AlkCβ Ig‐like domain. Regions contacting the DNA are highlighted in red.
Release of 3mA from methylated genomic DNA after a 5‐min incubation with either HCl, no enzyme (mock), PfAlkC (WT), or PfAlkC∆C (∆C). Values are mean ± SD (n = 3).
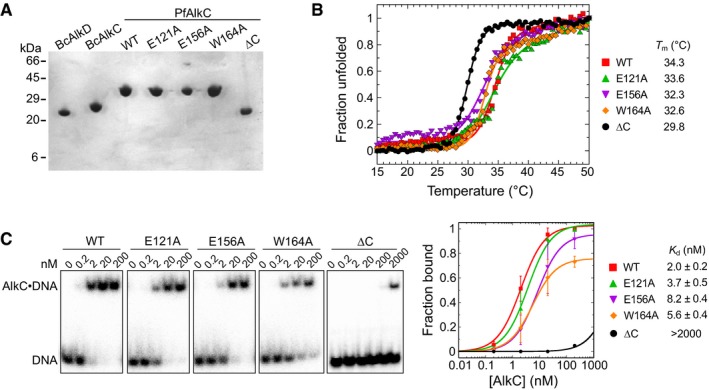
SDS–PAGE of purified proteins, stained with Coomassie Blue.
Thermal denaturation of wild‐type and PfAlkC mutants monitored by circular dichroism. The fraction of unfolded protein was expressed as the normalized molar ellipticity at 222 nm. Melting temperatures (T m) were derived by fitting the data to the equation (fraction unfolded) = 1/(1 + e(Tm−T)/k), where k denotes the cooperativity of the transition. Although PfAlkC∆C has a lower T m than the full‐length proteins, it is properly folded at 21°C at which biochemical assays were performed.
Electrophoretic mobility shift assay for PfAlkC mutants binding to 1 nM THF‐DNA of the sequence 32P‐d(GACCACTACACT(THF)ATTCCTAACAAC)/d(GTTGTTAGGAAT(T)AGTGTAGTGGTC) in 25 mM HEPES (pH 7.5), 50 mM KCl, 5 mM DTT, 5% glycerol, and 0.05 mg/ml BSA at 21°C for 30 min. Concentrations of PfAlkC are shown at the top of the representative gels. Electrophoretic separation was carried out on a Novex™ TBE gel (ThermoFisher Scientific). Quantitation is plotted on the right, in which each value is the mean ± SD (n = 3). Equilibrium dissociation constants (K d) were extracted by fitting the data to a two‐state binding model.
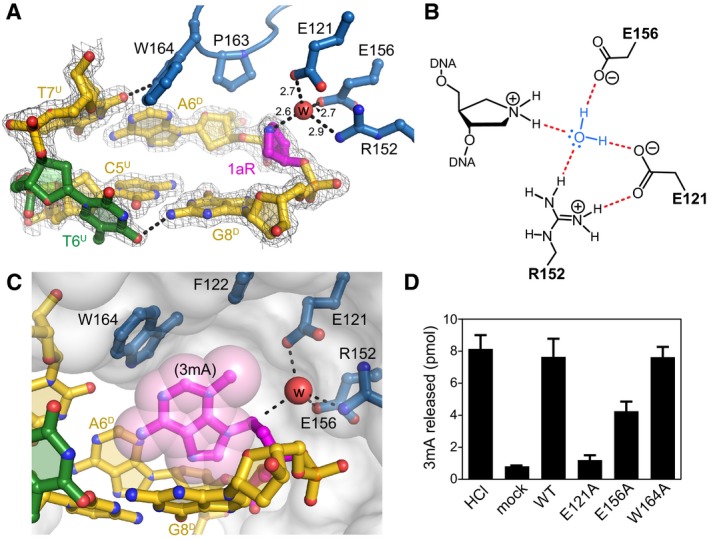
Close‐up view of the AlkC active site (blue) bound to 1aR‐DNA (gold). The 1aR and opposite thymine are magenta and green, respectively. Water is shown as a red sphere, and hydrogen bonds are depicted as dashed lines. Composite omit electron density contoured to 1σ is superimposed against only the DNA for clarity. Superscripts in nucleotide labels refer to the 1aR‐containing, damaged (D) strand or the opposite, undamaged (U) strand. The pentaerythritol propoxylate molecule that occupies the active site has been omitted for clarity (see Fig EV4B).
Schematic of the alignment of a catalytic water molecule (blue) against the 1aR oxocarbenium mimetic by AlkC active site residues. Hydrogen bonds are shown in red.
A hypothetical model for AlkC bound to a 3mA‐DNA substrate was generated by superimposing the 3mA deoxyribose ring onto that of 1aR in the crystal structure, followed by rotating about the 3mA χ (N‐glycosidic bond) torsion angle to maximize van der Waals interactions. The solvent accessible surface of AlkC is shown as a transparent white envelope.
Release of 3mA from methylated genomic DNA after a 5‐min incubation with either HCl, no enzyme (mock), PfAlkC (WT), E121A, E156A, or W164A. Values are mean ± SD (n = 3).
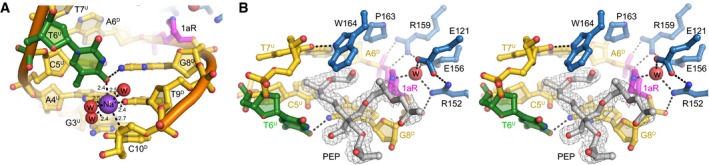
Na+ ion coordination in the 1aR‐DNA structure. Na+ and water oxygens are shown as purple and red spheres, respectively. 1aR is magenta, and the opposite thymidine T6 is green. Superscripts in nucleotide labels refer to the 1aR‐containing, damaged (D) strand or the opposite, undamaged (U) strand. Hydrogen bonds are shown as dashed lines with interatomic distances in Ångstroms.
Stereo‐views of the PfAlkC/1aR‐DNA active site. Active site residues are blue, and DNA is gold/magenta/green. Pentaerythritol propoxylate (PEP) sequestered from the solvent is in silver and superimposed against composite omit electron density contoured at 1σ. The putative catalytic water is shown as a red sphere. One arm of the PEP projects into the DNA kink between the 1aR and the flanking base pairs and thus may limit rotation of the 1aR ring back toward the DNA. The other two polymeric PEP arms project outward to solvent. The fourth PEP arm is not present; its terminal hydroxyl group forms a hydrogen bond to the displaced T6U thymine base. The position of the thymine is thus affected by the presence of the PEP in addition to its coordination by the Na+ ion. However, even in the absence of these stabilizing contacts, the T6U thymidine would be displaced into the minor groove as a result of the kink in the DNA. The space occupied by the PEP molecule would be occupied by the excised base both before and immediately after cleavage.
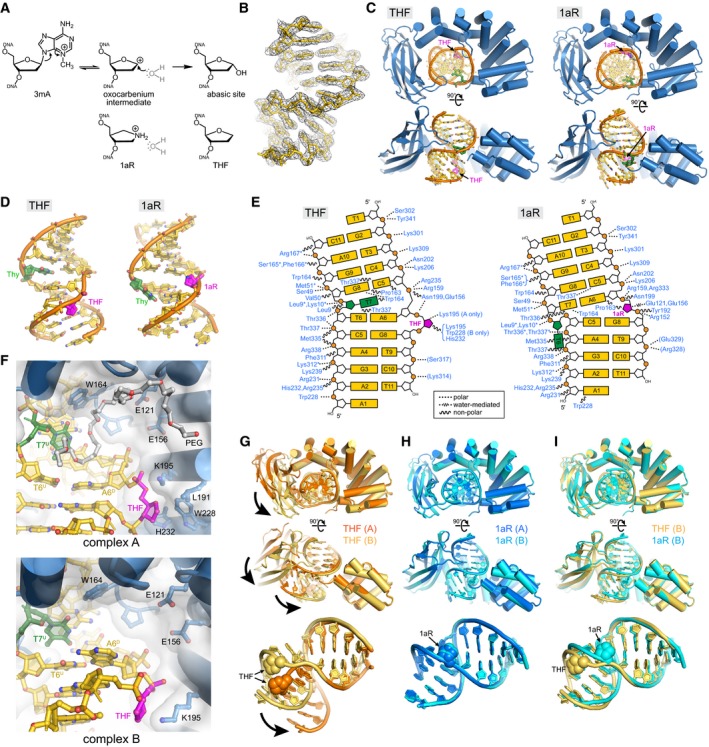
DNA glycosylase‐catalyzed reaction together with chemical structures of intermediate and product mimetics used in this study.
Annealed composite omit electron density contoured to 1σ is superposed onto the THF‐DNA crystallographic model.
Two views of the crystal structures, with protein in blue, DNA in gold, THF/1aR in magenta, and the thymidine opposite THF/1aR in green.
DNA structures extracted from the complexes.
Schematic of protein–DNA contacts. Contacts to the protein backbone are marked with an asterisk, and symmetry‐related contacts are in parentheses. PfAlkC maintains similar contacts with the DNA in THF and 1aR complexes relative to the position of the DNA bend and not to the position of the 1aR/THF.
Details of THF‐DNA bound outside of the PfAlkC active site. Both complexes in the crystallographic asymmetric unit are shown. A PEG 4,000 molecule (white carbons) fills the void in the catalytic pocket in complex A.
Superposition of the two protomers in the THF complex. Bold arrows highlight the difference in positions of the Ig‐like domain relative to the HLR domain and in the two DNA conformations.
Superposition of the two 1aR complexes.
Superposition of one protomer from each of the THF (gold) and 1aR (cyan) complexes.
- A
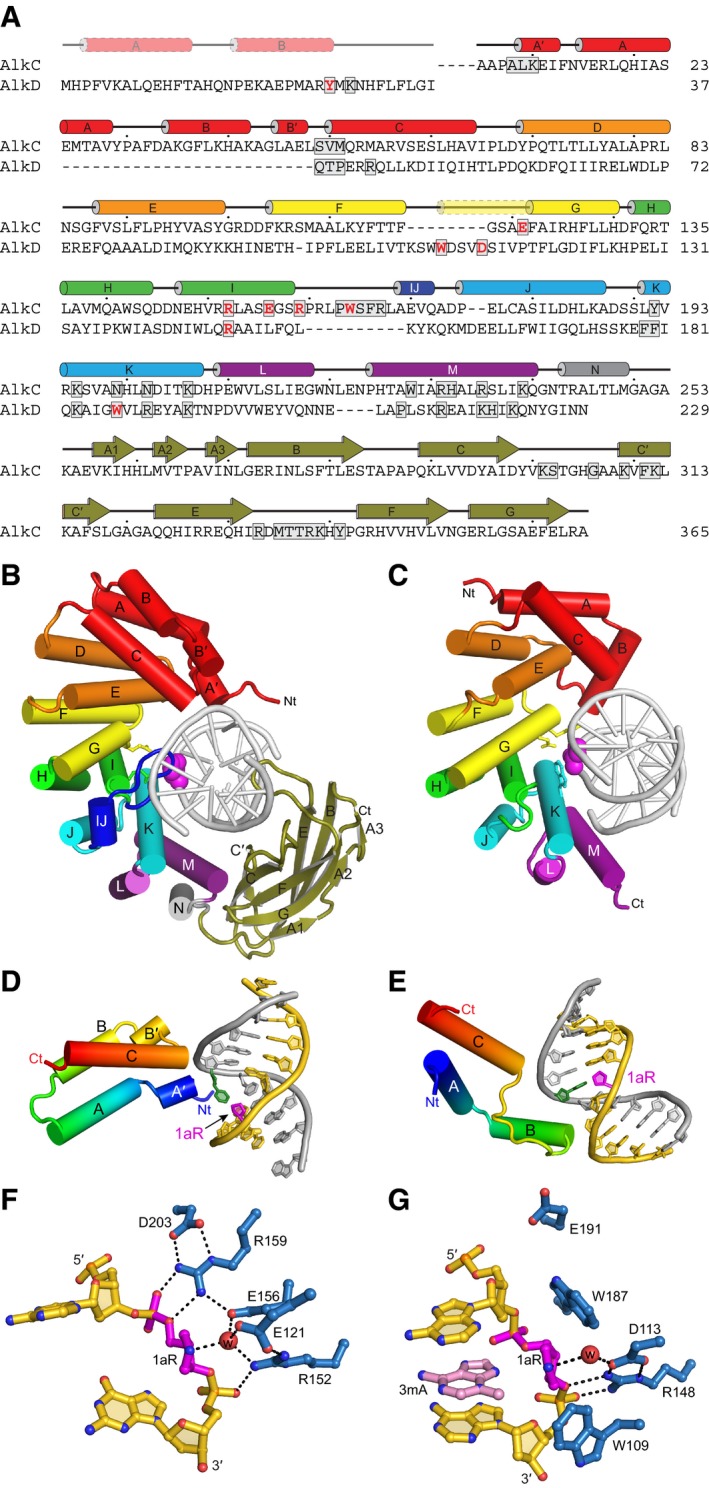
Structure‐based sequence alignment of PfAlkC and BcAlkD. Active site residues are red and DNA‐interacting residues are boxed. Secondary structures derived from the crystal structures are shown above the sequences. The transparent N‐terminal segment of helix αG (yellow) is unique to AlkD and helix αIJ (dark blue) is unique to AlkC.
- B, C
Crystal structures of 1aR‐DNA complexes of PfAlkC (B) and BcAlkD (C, PDB ID 5CLD). The N‐terminal helical bundles are red; HEAT‐like repeats are orange, yellow, green, cyan, and purple; and the Ig‐like domain of AlkC is olive. Side chains of active site residues are shown as sticks, and the 1aR moieties are shown as magenta spheres.
- D, E
Interactions between the N‐terminal helical bundles of PfAlkC (D) and BcAlkD (E) and DNA. Protein is colored rainbow from N‐ (blue) to C‐terminus (red). DNA strands are colored gold and silver, and 1aR and opposite thymine are magenta and green, respectively.
- F, G
Active sites of PfAlkC (F) and BcAlkD (G). Protein residues are colored blue, DNA gold, 1aR magenta, and 3mA nucleobase pink. The catalytic water is shown as a red sphere, and hydrogen bonds are depicted as dashed lines.
- A
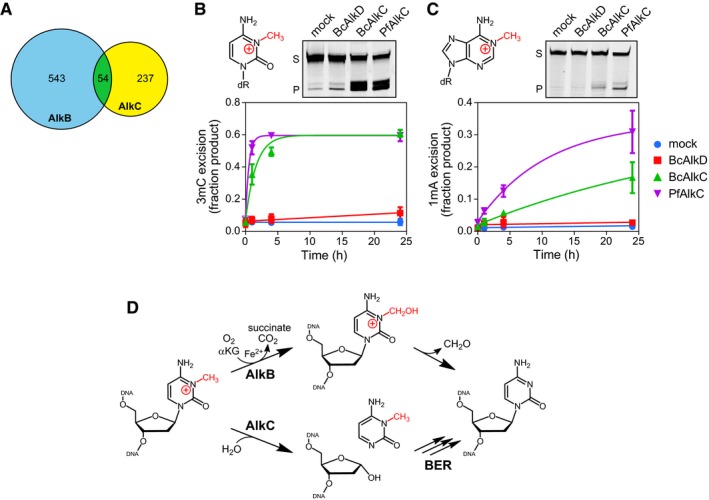
Venn diagram of the numbers of bacterial species containing either AlkB (blue), AlkC (yellow), or both (green).
- B, C
Chemical structures and in vitro base excision of 3mC (B) and 1mA (C) from 25‐mer double‐stranded oligodeoxyribonucleotides. Representative denaturing electrophoresis gels show substrate (S) and product (P) after a 24‐h incubation with either no enzyme (mock), BcAlkD, BcAlkC (AlkCα), or PfAlkC (AlkCβ). Plots show quantified time courses from three experiments (values are mean ± SD).
- D
3mC may be repaired in bacteria by either AlkB‐catalyzed oxidative demethylation or AlkC‐catalyzed base excision. αKG, α‐ketoglutarate.
References
-
- Backe PH, Simm R, Laerdahl JK, Dalhus B, Fagerlund A, Okstad OA, Rognes T, Alseth I, Kolsto AB, Bjoras M (2013) A new family of proteins related to the HEAT‐like repeat DNA glycosylases with affinity for branched DNA structures. J Struct Biol 183: 66–75 - PubMed
-
- Becker S, Groner B, Muller CW (1998) Three‐dimensional structure of the Stat3β homodimer bound to DNA. Nature 394: 145 - PubMed
-
- Birkeland NK, Anensen H, Knaevelsrud I, Kristoffersen W, Bjoras M, Robb FT, Klungland A, Bjelland S (2002) Methylpurine DNA glycosylase of the hyperthermophilic archaeon Archaeoglobus fulgidus . Biochemistry 41: 12697–12705 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
