

The Molecular Basis of Noncanonical Bacterial Morphology
- PMID: 29056293
- PMCID: PMC5834356
- DOI: 10.1016/j.tim.2017.09.012
The Molecular Basis of Noncanonical Bacterial Morphology
Abstract
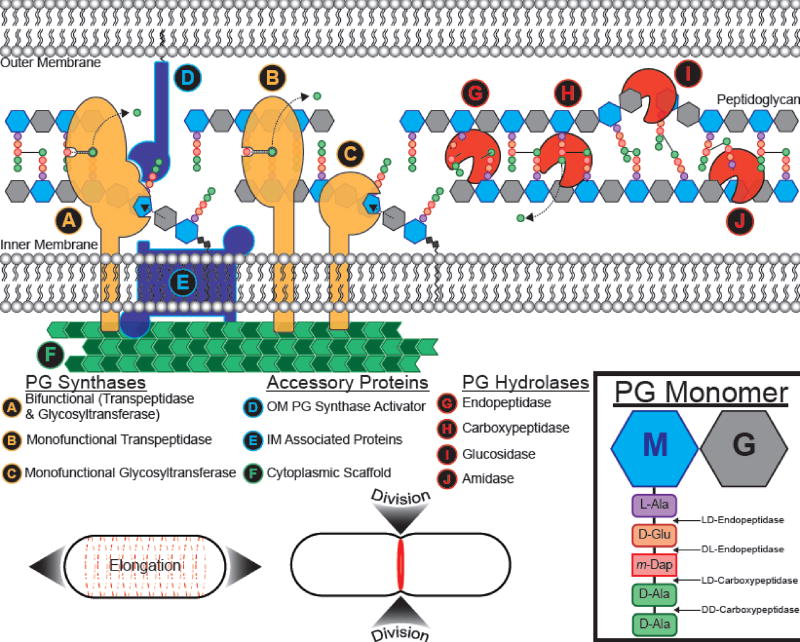
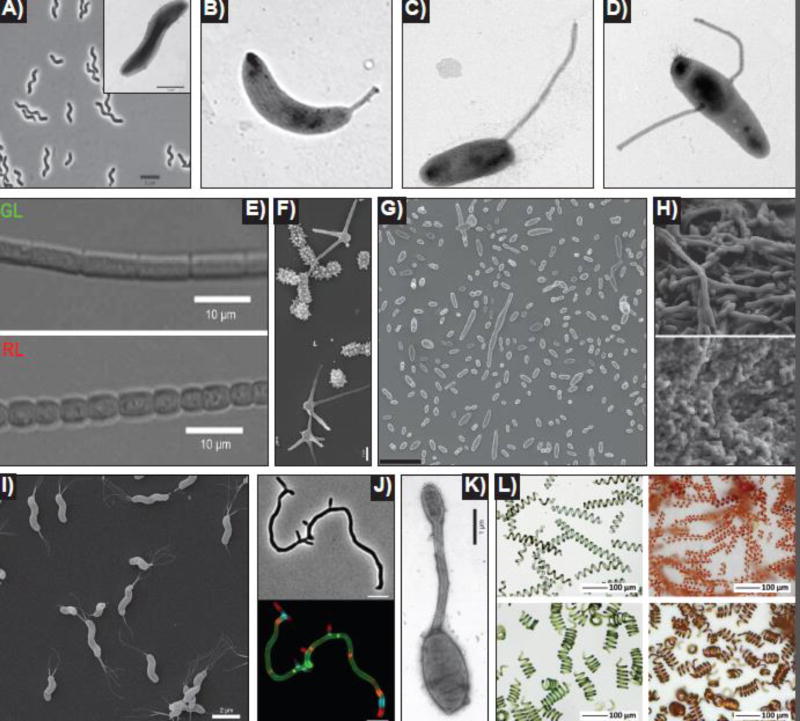
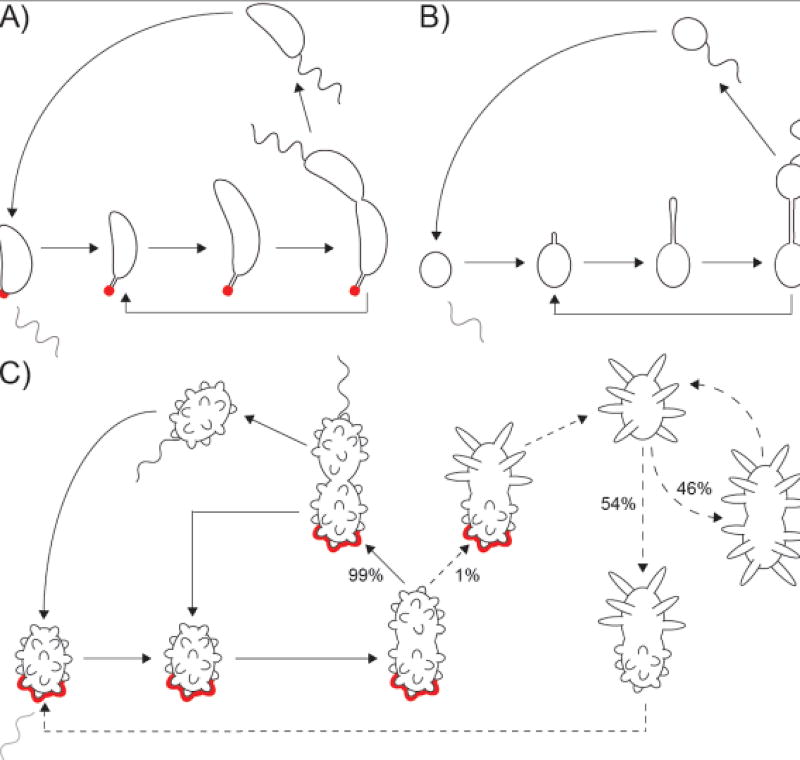
Bacteria come in a wide variety of shapes and sizes. The true picture of bacterial morphological diversity is likely skewed due to an experimental focus on pathogens and industrially relevant organisms. Indeed, most of the work elucidating the genes and molecular processes involved in maintaining bacterial morphology has been limited to rod- or coccal-shaped model systems. The mechanisms of shape evolution, the molecular processes underlying diverse shapes and growth modes, and how individual cells can dynamically modulate their shape are just beginning to be revealed. Here we discuss recent work aimed at advancing our knowledge of shape diversity and uncovering the molecular basis for shape generation in noncanonical and morphologically complex bacteria.
Keywords: bacterial shape; morphological engineering; morphology; peptidoglycan; pleomorphism.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Allan EJ, et al. Bacterial L-forms. Adv Appl Microbiol. 2009;68:1–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
