

Early-Life Gene Expression in Neurons Modulates Lasting Epigenetic States
- PMID: 29056337
- PMCID: PMC5693680
- DOI: 10.1016/j.cell.2017.09.047
Early-Life Gene Expression in Neurons Modulates Lasting Epigenetic States
Abstract
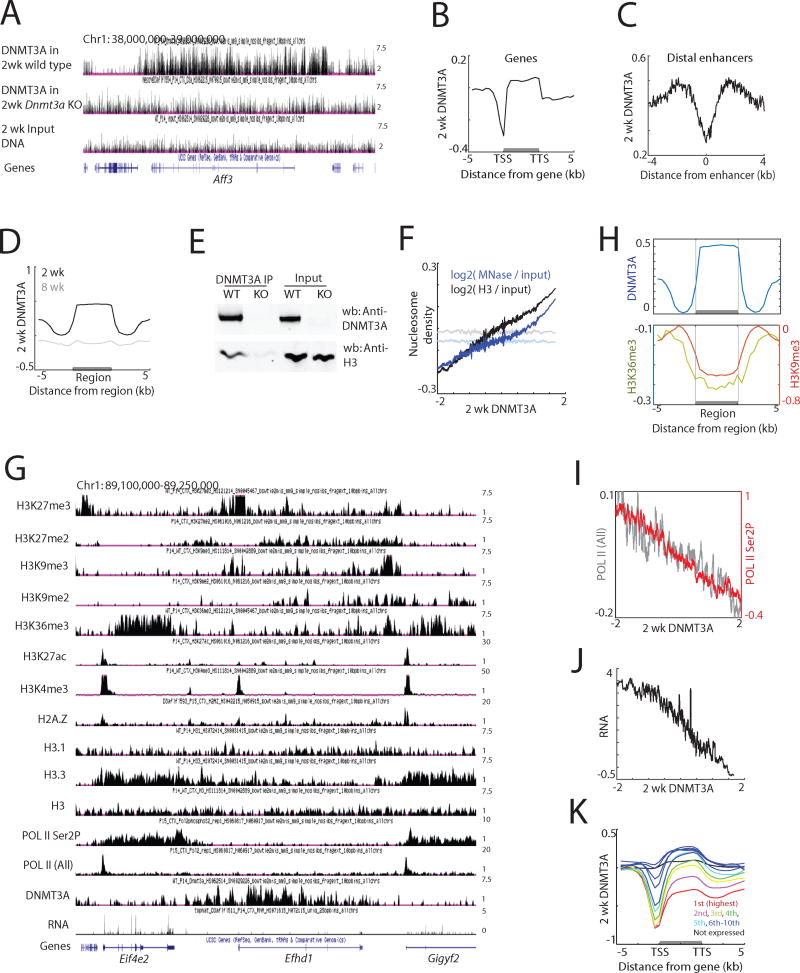
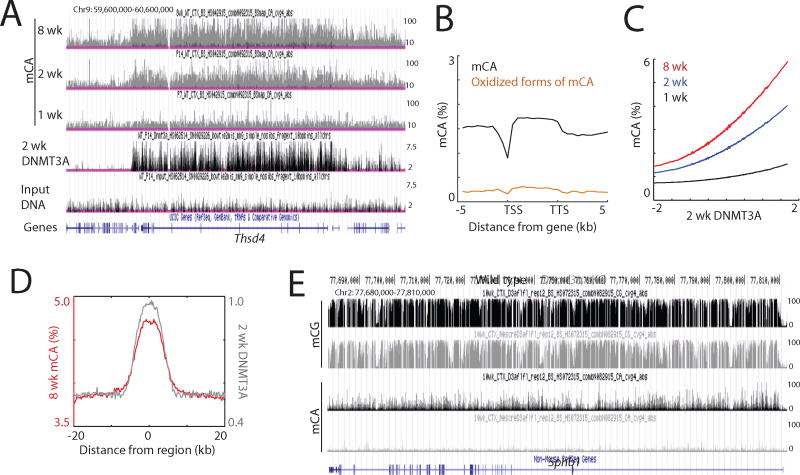
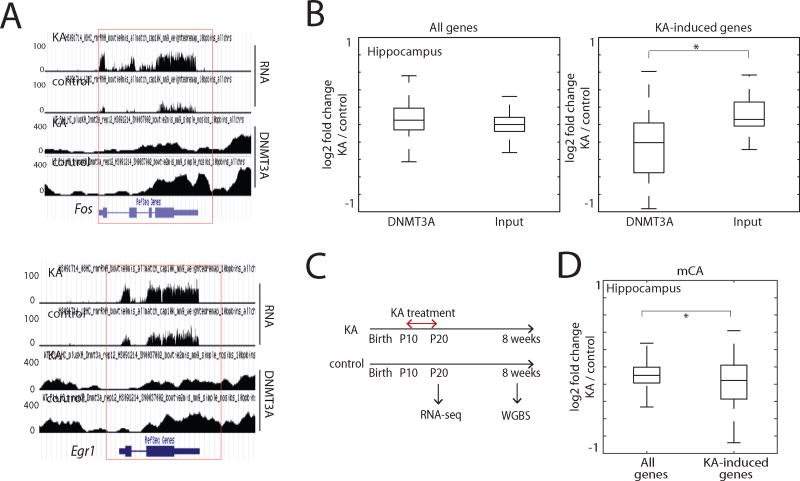
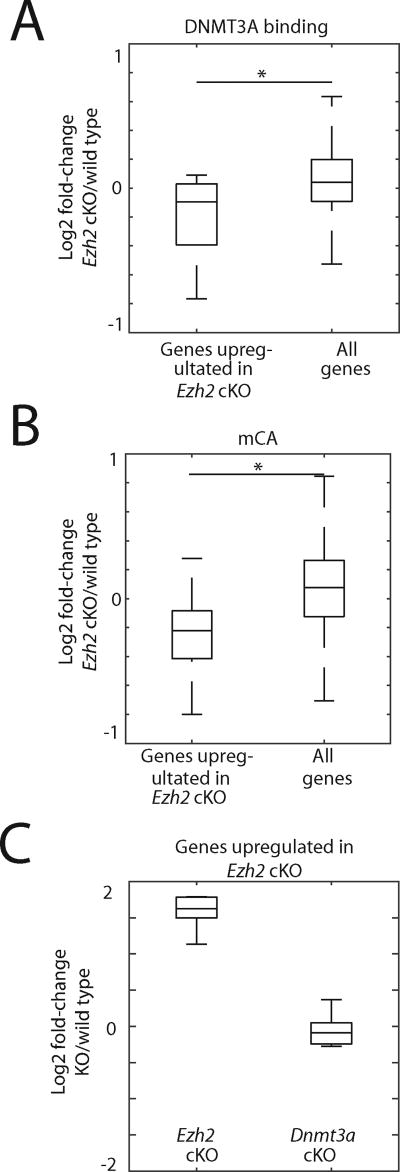
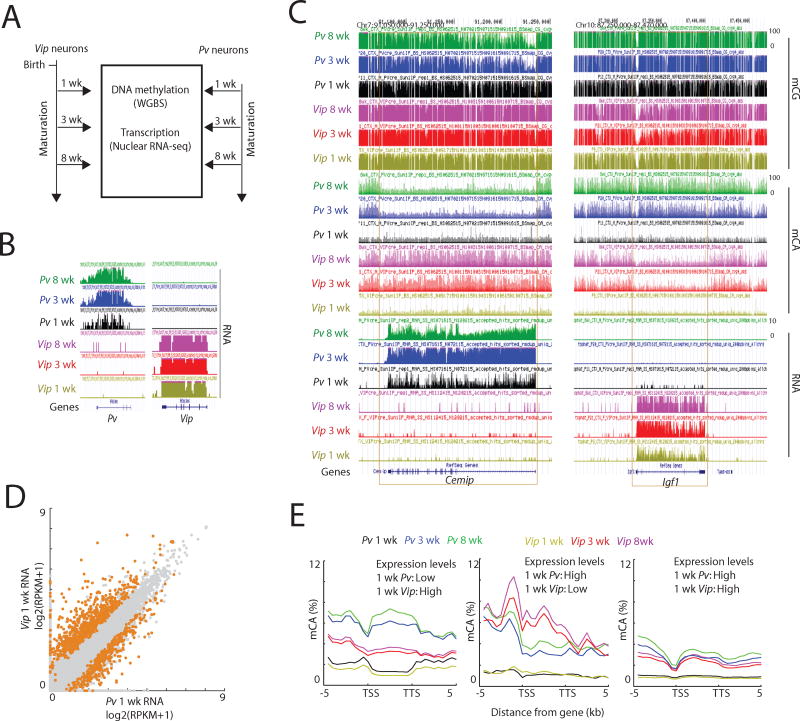
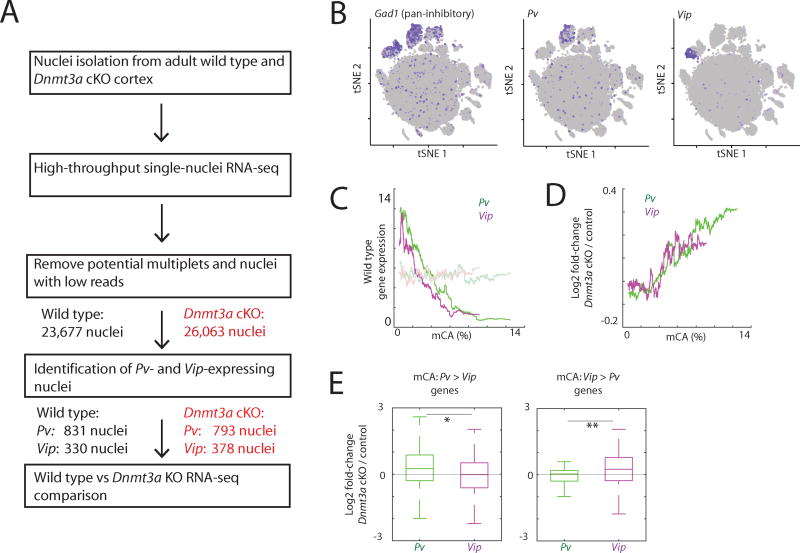
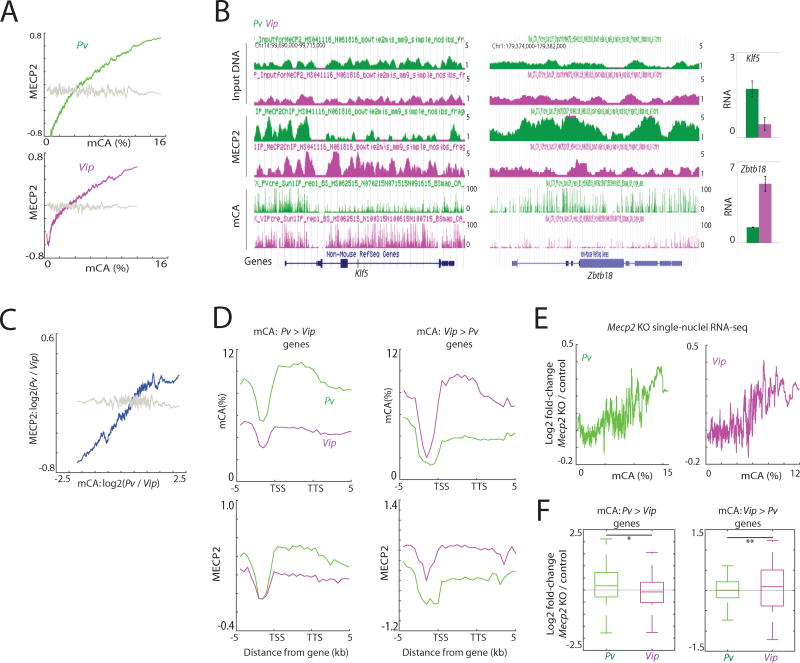
In mammals, the environment plays a critical role in promoting the final steps in neuronal development during the early postnatal period. While epigenetic factors are thought to contribute to this process, the underlying molecular mechanisms remain poorly understood. Here, we show that in the brain during early life, the DNA methyltransferase DNMT3A transiently binds across transcribed regions of lowly expressed genes, and its binding specifies the pattern of DNA methylation at CA sequences (mCA) within these genes. We find that DNMT3A occupancy and mCA deposition within the transcribed regions of genes is negatively regulated by gene transcription and may be modified by early-life experience. Once deposited, mCA is bound by the methyl-DNA-binding protein MECP2 and functions in a rheostat-like manner to fine-tune the cell-type-specific transcription of genes that are critical for brain function.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Epigenetics: Leaving a lasting mark.Nat Rev Neurosci. 2017 Dec;18(12):710. doi: 10.1038/nrn.2017.144. Epub 2017 Nov 9. Nat Rev Neurosci. 2017. PMID: 29118448 No abstract available.
References
-
- Baubec T, Colombo DF, Wirbelauer C, Schmidt J, Burger L, Krebs AR, Akalin A, Schubeler D. Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature 2015 - PubMed
-
- Baubec T, Ivanek R, Lienert F, Schubeler D. Methylation-dependent and -independent genomic targeting principles of the MBD protein family. Cell. 2013;153:480–492. - PubMed
-
- Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002;16:6–21. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
