

Selective Inhibition of FOXO1 Activator/Repressor Balance Modulates Hepatic Glucose Handling
- PMID: 29056338
- PMCID: PMC5687849
- DOI: 10.1016/j.cell.2017.09.045
Selective Inhibition of FOXO1 Activator/Repressor Balance Modulates Hepatic Glucose Handling
Abstract
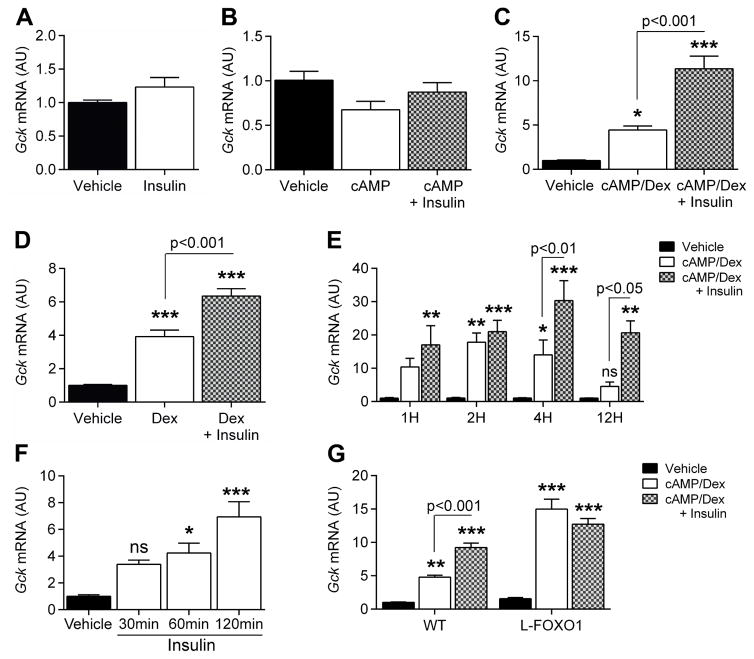
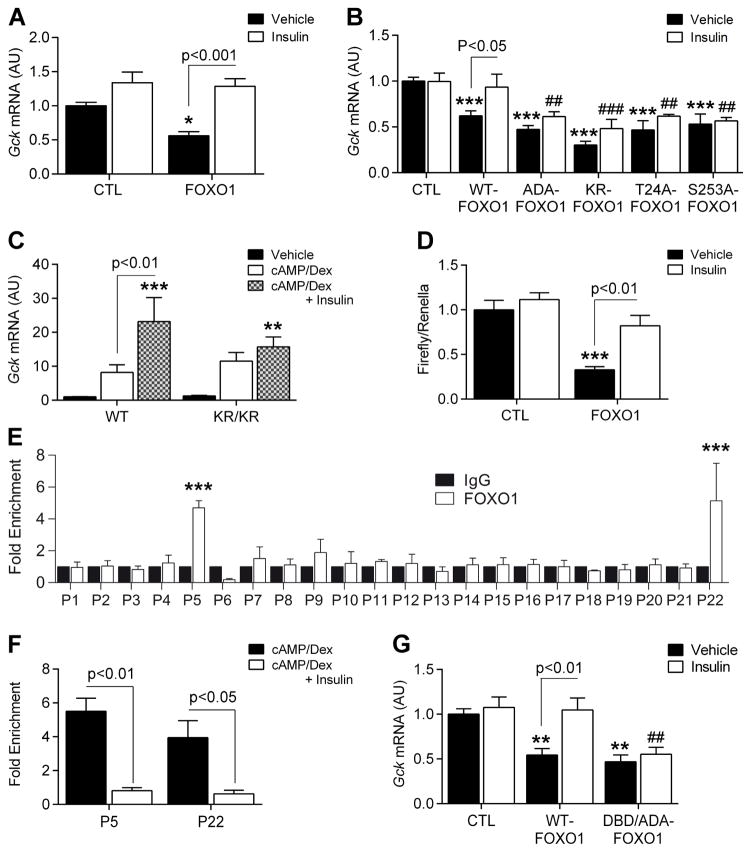
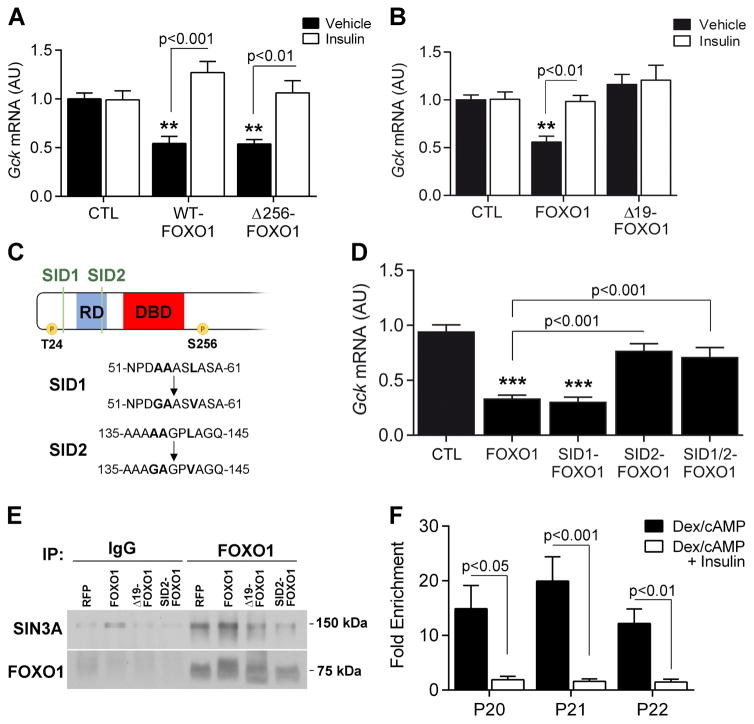
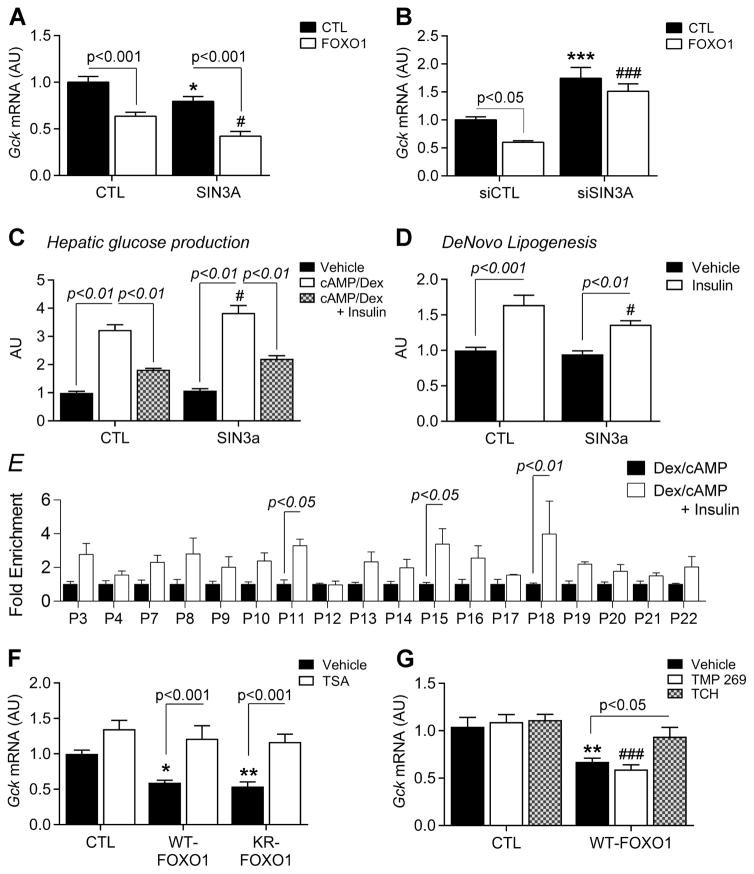
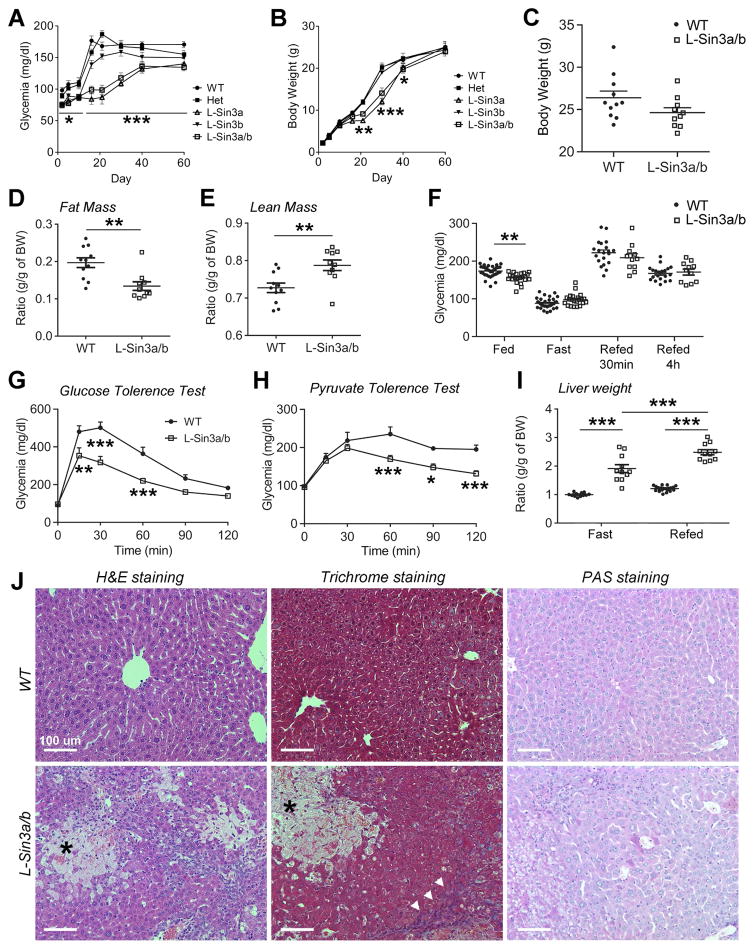
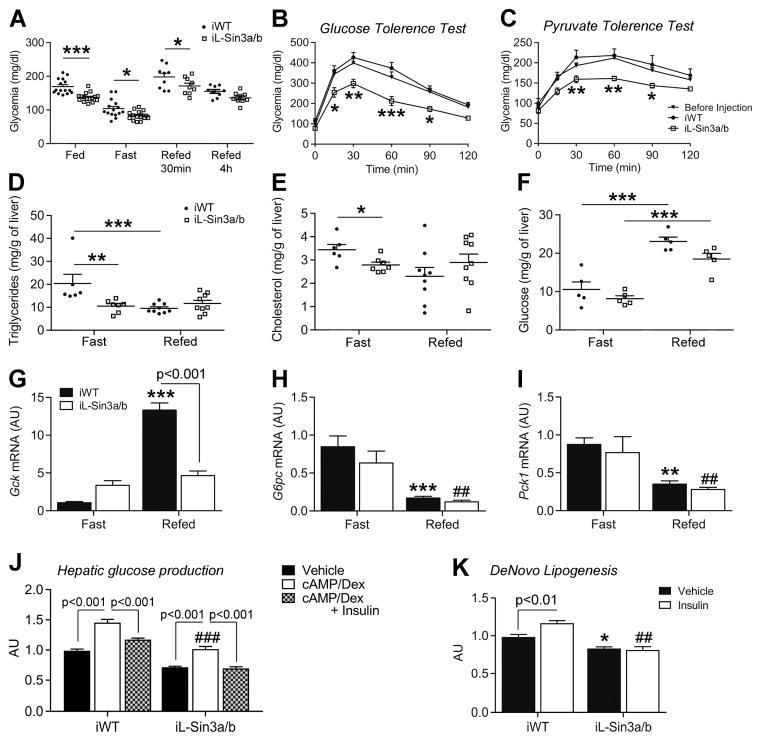
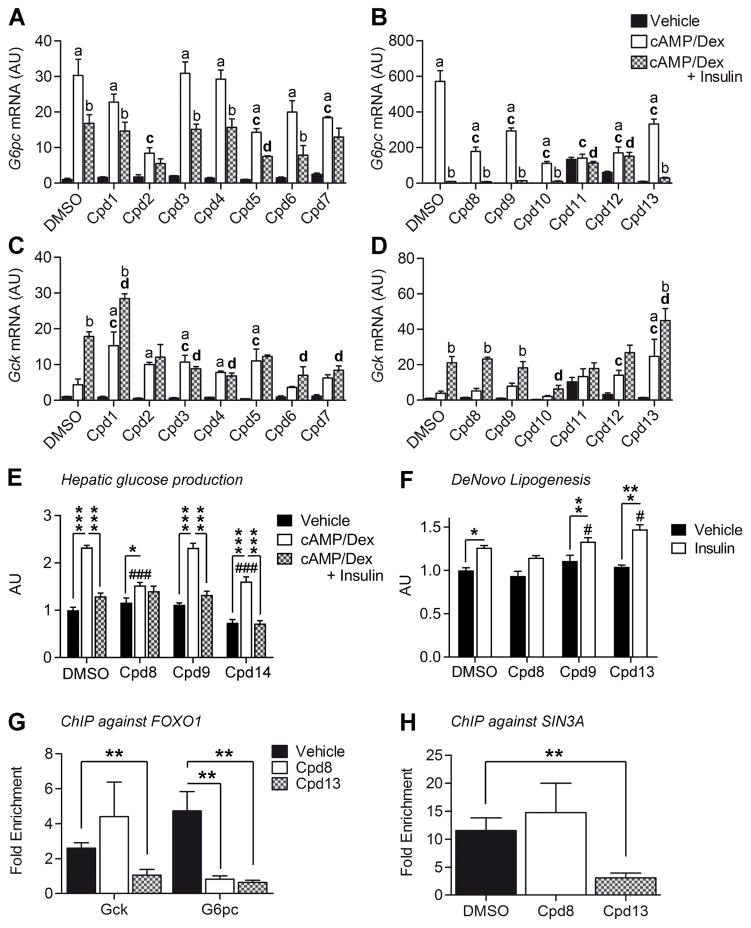
Insulin resistance is a hallmark of diabetes and an unmet clinical need. Insulin inhibits hepatic glucose production and promotes lipogenesis by suppressing FOXO1-dependent activation of G6pase and inhibition of glucokinase, respectively. The tight coupling of these events poses a dual conundrum: mechanistically, as the FOXO1 corepressor of glucokinase is unknown, and clinically, as inhibition of glucose production is predicted to increase lipogenesis. Here, we report that SIN3A is the insulin-sensitive FOXO1 corepressor of glucokinase. Genetic ablation of SIN3A abolishes nutrient regulation of glucokinase without affecting other FOXO1 target genes and lowers glycemia without concurrent steatosis. To extend this work, we executed a small-molecule screen and discovered selective inhibitors of FOXO-dependent glucose production devoid of lipogenic activity in hepatocytes. In addition to identifying a novel mode of insulin action, these data raise the possibility of developing selective modulators of unliganded transcription factors to dial out adverse effects of insulin sensitizers.
Keywords: diabetes; drug therapy; hepatic glucose production; hepatosteatosis; insulin resistance; insulin sensitizers; lipogenesis; selective modulators; small molecule inhibitor; transcription repressor.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Diabetes: A new hope for insulin-sensitizing drugs.Nat Rev Endocrinol. 2017 Nov 9;13(12):687. doi: 10.1038/nrendo.2017.150. Nat Rev Endocrinol. 2017. PMID: 29118351 No abstract available.
References
-
- Altomonte J, Richter A, Harbaran S, Suriawinata J, Nakae J, Thung SN, Meseck M, Accili D, Dong H. Inhibition of Foxo1 function is associated with improved fasting glycemia in diabetic mice. Am J Physiol - Endocrinol Metab. 2003;285:E718–E728. - PubMed
-
- Berkowitz K, Peters R, Kjos SL, Goico J, Marroquin A, Dunn ME, Xiang A, Azen S, Buchanan TA. Effect of troglitazone on insulin sensitivity and pancreatic beta-cell function in women at high risk for NIDDM. Diabetes. 1996;45:1572–1579. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
