

Two Disease-Causing SNAP-25B Mutations Selectively Impair SNARE C-terminal Assembly
- PMID: 29056461
- PMCID: PMC5805579
- DOI: 10.1016/j.jmb.2017.10.012
Two Disease-Causing SNAP-25B Mutations Selectively Impair SNARE C-terminal Assembly
Abstract
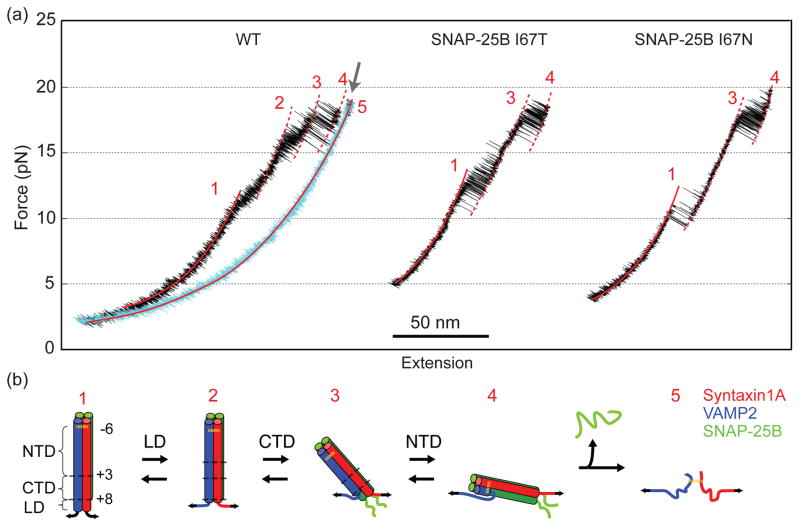
Synaptic exocytosis relies on assembly of three soluble N-ethylmaleimide-sensitive factor attachment receptor (SNARE) proteins into a parallel four-helix bundle to drive membrane fusion. SNARE assembly occurs by stepwise zippering of the vesicle-associated SNARE (v-SNARE) onto a binary SNARE complex on the target plasma membrane (t-SNARE). Zippering begins with slow N-terminal association followed by rapid C-terminal zippering, which serves as a power stroke to drive membrane fusion. SNARE mutations have been associated with numerous diseases, especially neurological disorders. It remains unclear how these mutations affect SNARE zippering, partly due to difficulties to quantify the energetics and kinetics of SNARE assembly. Here, we used single-molecule optical tweezers to measure the assembly energy and kinetics of SNARE complexes containing single mutations I67T/N in neuronal SNARE synaptosomal-associated protein of 25kDa (SNAP-25B), which disrupt neurotransmitter release and have been implicated in neurological disorders. We found that both mutations significantly reduced the energy of C-terminal zippering by ~10 kBT, but did not affect N-terminal assembly. In addition, we observed that both mutations lead to unfolding of the C-terminal region in the t-SNARE complex. Our findings suggest that both SNAP-25B mutations impair synaptic exocytosis by destabilizing SNARE assembly, rather than stabilizing SNARE assembly as previously proposed. Therefore, our measurements provide insights into the molecular mechanism of the disease caused by SNARE mutations.
Keywords: SNARE assembly; membrane fusion; neuropathy; optical tweezers; protein folding.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures





References
-
- Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M, Parlati F, et al. SNAREpins: Minimal machinery for membrane fusion. Cell. 1998;92:759–72. - PubMed
-
- Sollner T, Whiteheart SW, Brunner M, Erdjument-Bromage H, Geromanos S, Tempst P, et al. SNAP receptors implicated in vesicle targeting and fusion. Nature. 1993;362:318–24. - PubMed
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin - a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
