

Editing inducer elements increases A-to-I editing efficiency in the mammalian transcriptome
- PMID: 29061182
- PMCID: PMC5654063
- DOI: 10.1186/s13059-017-1324-x
Editing inducer elements increases A-to-I editing efficiency in the mammalian transcriptome
Abstract
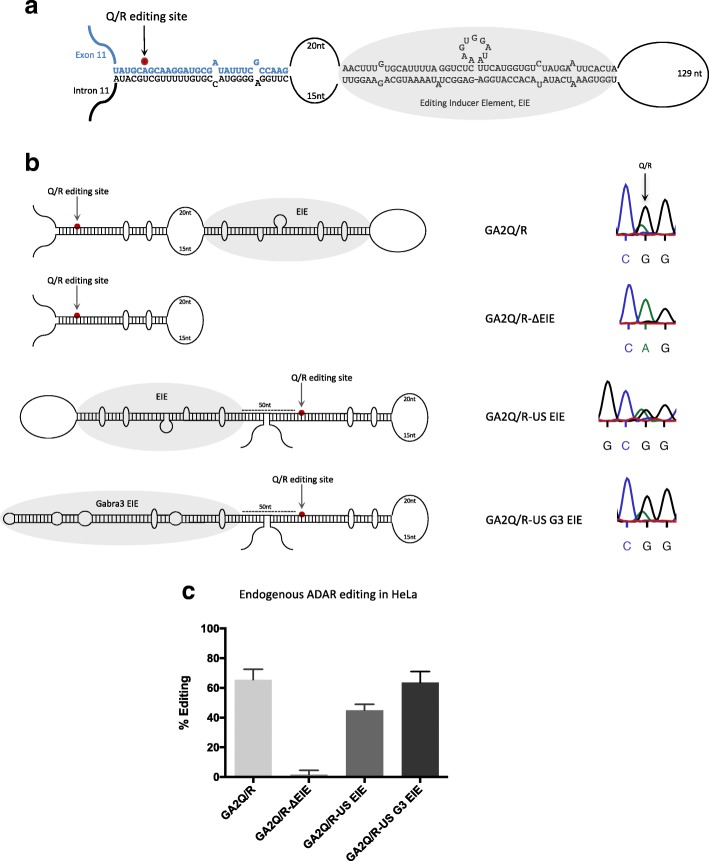
Background: Adenosine to inosine (A-to-I) RNA editing has been shown to be an essential event that plays a significant role in neuronal function, as well as innate immunity, in mammals. It requires a structure that is largely double-stranded for catalysis but little is known about what determines editing efficiency and specificity in vivo. We have previously shown that some editing sites require adjacent long stem loop structures acting as editing inducer elements (EIEs) for efficient editing.
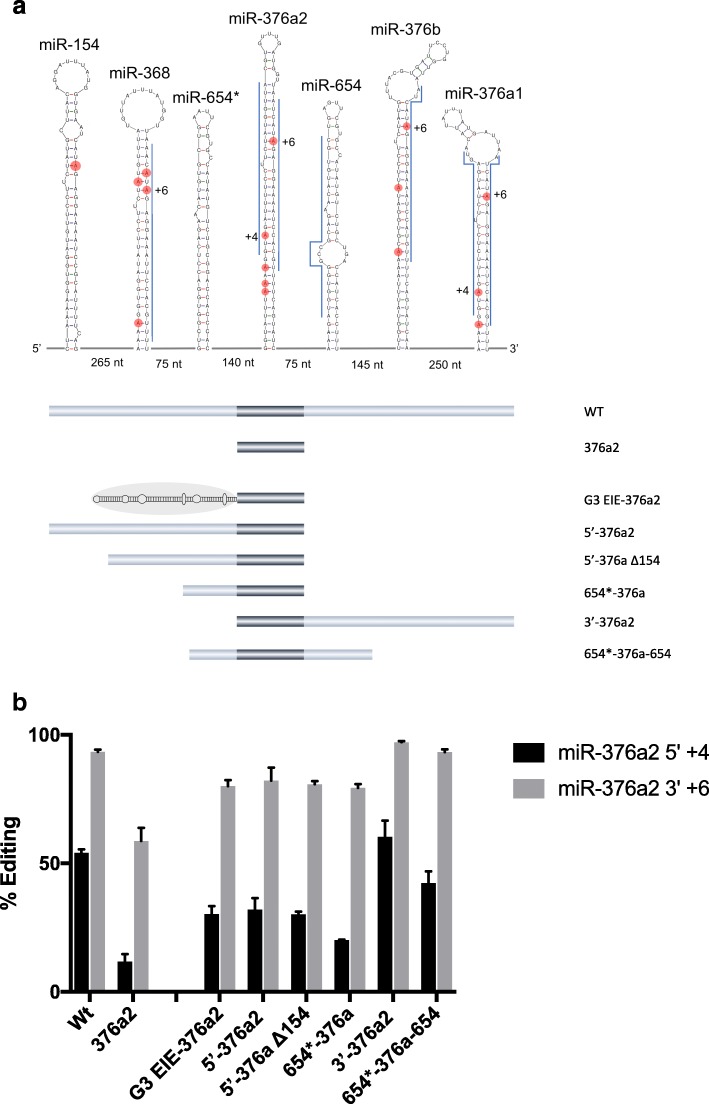
Results: The glutamate receptor subunit A2 is edited at the Q/R site in almost 100% of all transcripts. We show that efficient editing at the Q/R site requires an EIE in the downstream intron, separated by an internal loop. Also, other efficiently edited sites are flanked by conserved, highly structured EIEs and we propose that this is a general requisite for efficient editing, while sites with low levels of editing lack EIEs. This phenomenon is not limited to mRNA, as non-coding primary miRNAs also use EIEs to recruit ADAR to specific sites.
Conclusions: We propose a model where two regions of dsRNA are required for efficient editing: first, an RNA stem that recruits ADAR and increases the local concentration of the enzyme, then a shorter, less stable duplex that is ideal for efficient and specific catalysis. This discovery changes the way we define and determine a substrate for A-to-I editing. This will be important in the discovery of novel editing sites, as well as explaining cases of altered editing in relation to disease.
Keywords: ADAR; Adenosine deamination; EIE; Glutamate receptor; RNA editing; miRNA.
Conflict of interest statement
Ethics approval
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures





Comment in
-
Promoting RNA editing by ADAR attraction.Genome Biol. 2017 Oct 23;18(1):196. doi: 10.1186/s13059-017-1343-7. Genome Biol. 2017. PMID: 29061157 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
