

NLRP3 Is a Critical Regulator of Inflammation and Innate Immune Cell Response during Mycoplasma pneumoniae Infection
- PMID: 29061706
- PMCID: PMC5736809
- DOI: 10.1128/IAI.00548-17
NLRP3 Is a Critical Regulator of Inflammation and Innate Immune Cell Response during Mycoplasma pneumoniae Infection
Abstract
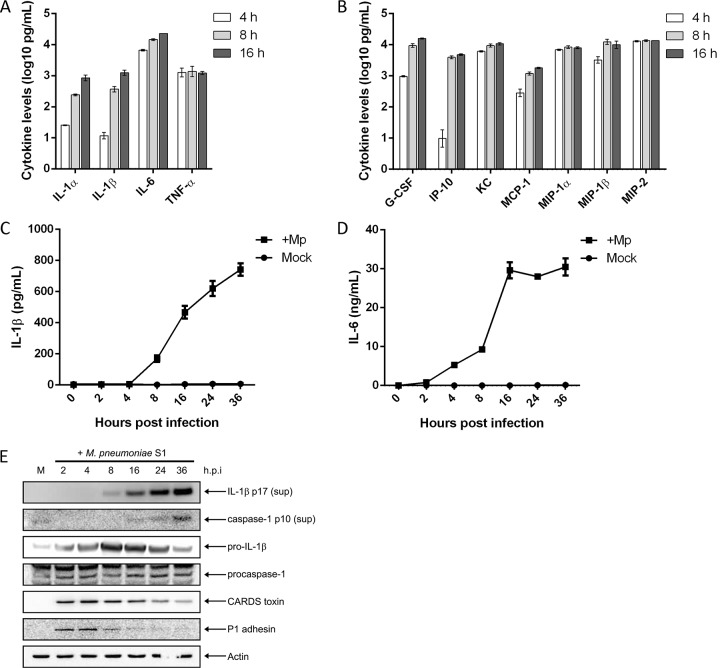
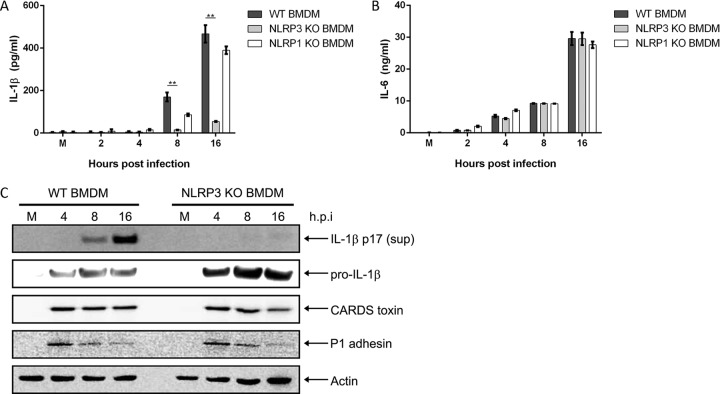
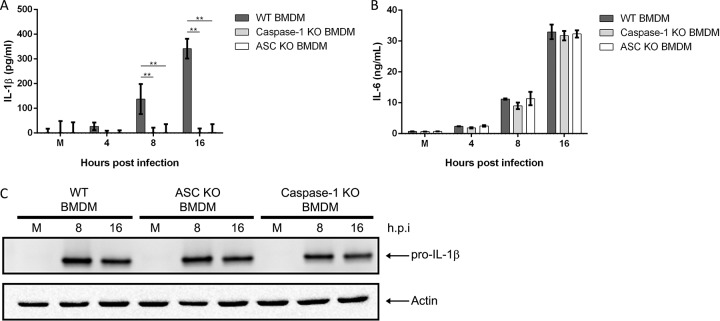
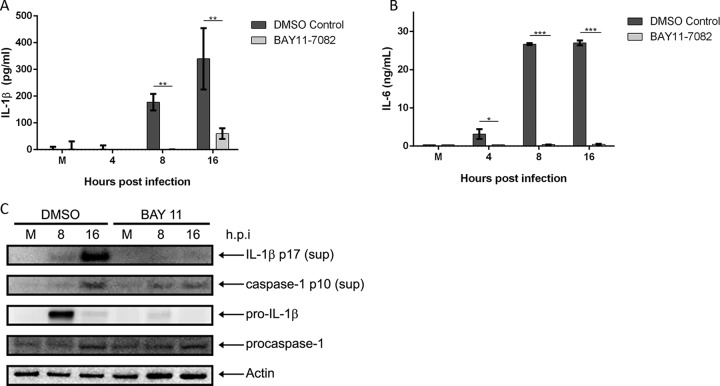
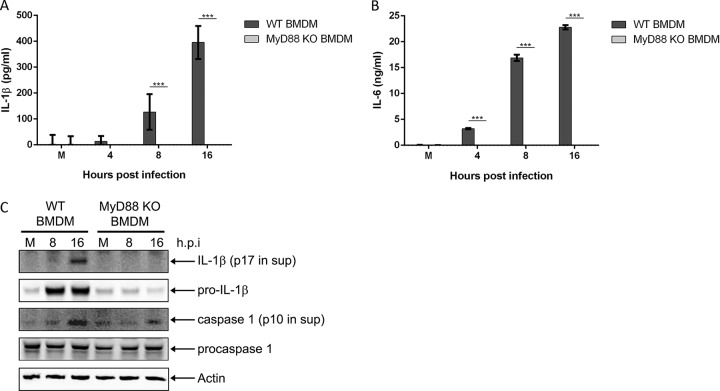
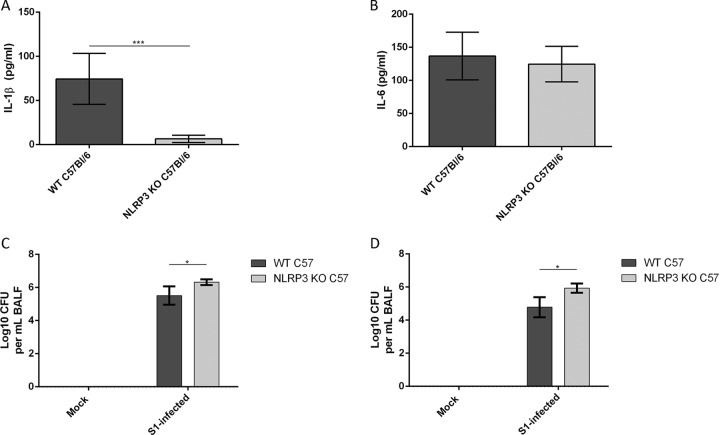
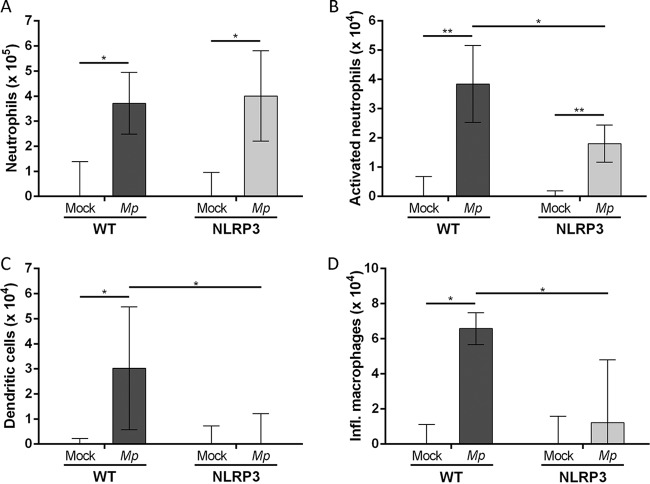
Mycoplasma pneumoniae is an atypical bacterial respiratory pathogen known to cause a range of airway inflammation and lung and extrapulmonary pathologies. We recently reported that an M. pneumoniae-derived ADP-ribosylating and vacuolating toxin called community-acquired respiratory distress syndrome (CARDS) toxin is capable of triggering NLRP3 (NLR-family, leucine-rich repeat protein 3) inflammasome activation and interleukin-1β (IL-1β) secretion in macrophages. However, it is unclear whether the NLRP3 inflammasome is important for the immune response during M. pneumoniae acute infection. In the current study, we utilized in vitro and in vivo models of M. pneumoniae infection to characterize the role of the NLRP3 inflammasome during acute infection. M. pneumoniae-infected macrophages deficient for inflammasome components NLRP3, ASC (apoptosis speck-like protein containing a caspase activation and recruitment domain), or caspase-1 failed to process and secrete IL-1β. The MyD88/NF-κB signaling pathway was found to be critical for proinflammatory gene expression in macrophages infected with M. pneumoniae C57BL/6 mice deficient for NLRP3 expression were unable to produce IL-1β in the airways during acute infection, and lack of this inflammatory response led to deficient immune cell activation and delayed bacterial clearance. These findings are the first to report the importance of the NLRP3 inflammasome in regulating the inflammatory response and influencing the progression of M. pneumoniae during acute infection.
Keywords: ADP-ribosylation; CARDS toxin; Mycoplasma pneumoniae; NLRP3; inflammasome; interleukin-1β.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Jain S, Williams DJ, Arnold SR, Ampofo K, Bramley AM, Reed C, Stockmann C, Anderson EJ, Grijalva CG, Self WH, Zhu Y, Patel A, Hymas W, Chappell JD, Kaufman RA, Kan JH, Dansie D, Lenny N, Hillyard DR, Haynes LM, Levine M, Lindstrom S, Winchell JM, Katz JM, Erdman D, Schneider E, Hicks LA, Wunderink RG, Edwards KM, Pavia AT, McCullers JA, Finelli L, Team CES. 2015. Community-acquired pneumonia requiring hospitalization among U.S. children. N Engl J Med 372:835–845. doi: 10.1056/NEJMoa1405870. - DOI - PMC - PubMed
-
- Jain S, Self WH, Wunderink RG, Fakhran S, Balk R, Bramley AM, Reed C, Grijalva CG, Anderson EJ, Courtney DM, Chappell JD, Qi C, Hart EM, Carroll F, Trabue C, Donnelly HK, Williams DJ, Zhu Y, Arnold SR, Ampofo K, Waterer GW, Levine M, Lindstrom S, Winchell JM, Katz JM, Erdman D, Schneider E, Hicks LA, McCullers JA, Pavia AT, Edwards KM, Finelli L, CDC EPIC Study Team. 2015. Community-acquired pneumonia requiring hospitalization among U.S. adults. N Engl J Med 373:415–427. doi: 10.1056/NEJMoa1500245. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
