

Geographic mosaics and changing rates of cereal domestication
- PMID: 29061901
- PMCID: PMC5665816
- DOI: 10.1098/rstb.2016.0429
Geographic mosaics and changing rates of cereal domestication
Abstract
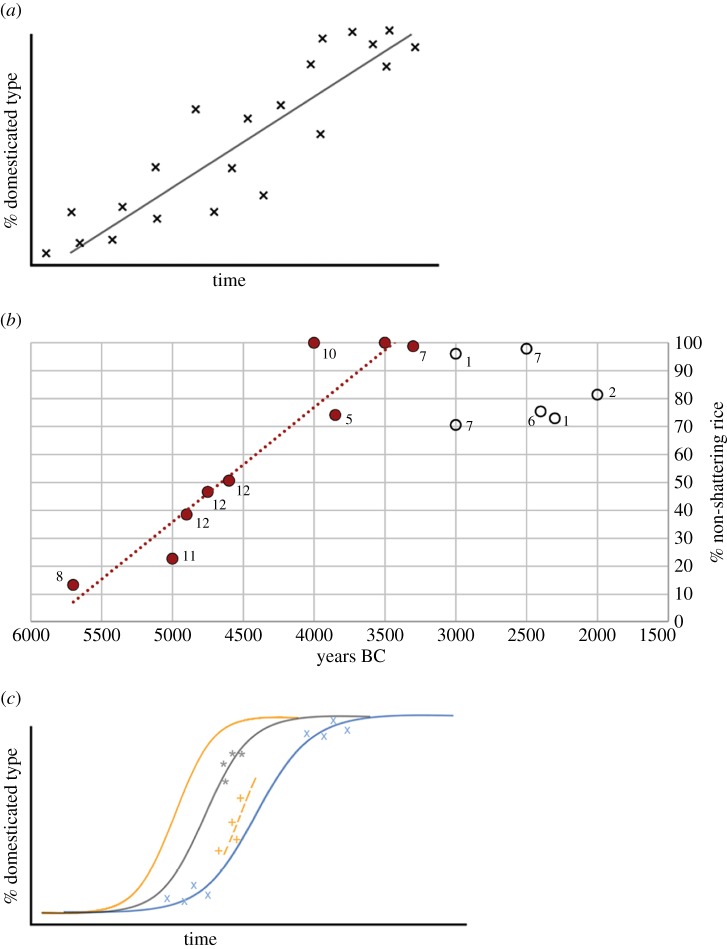
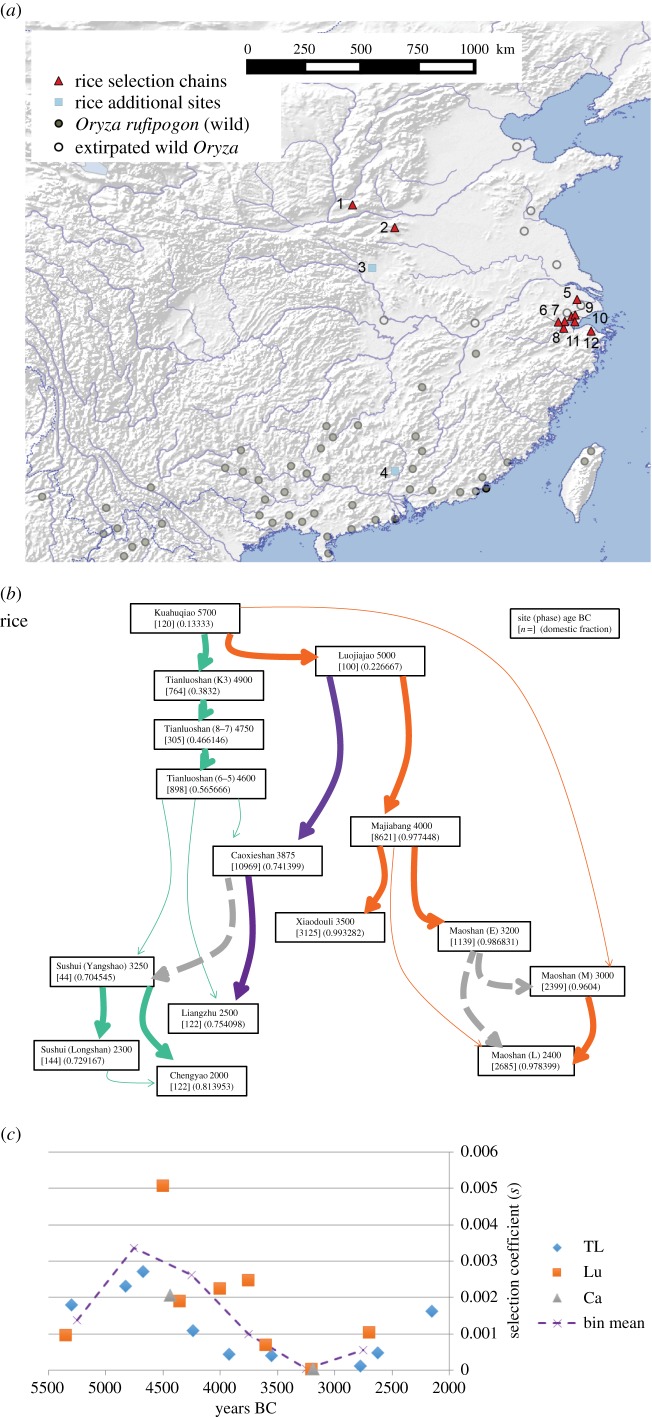
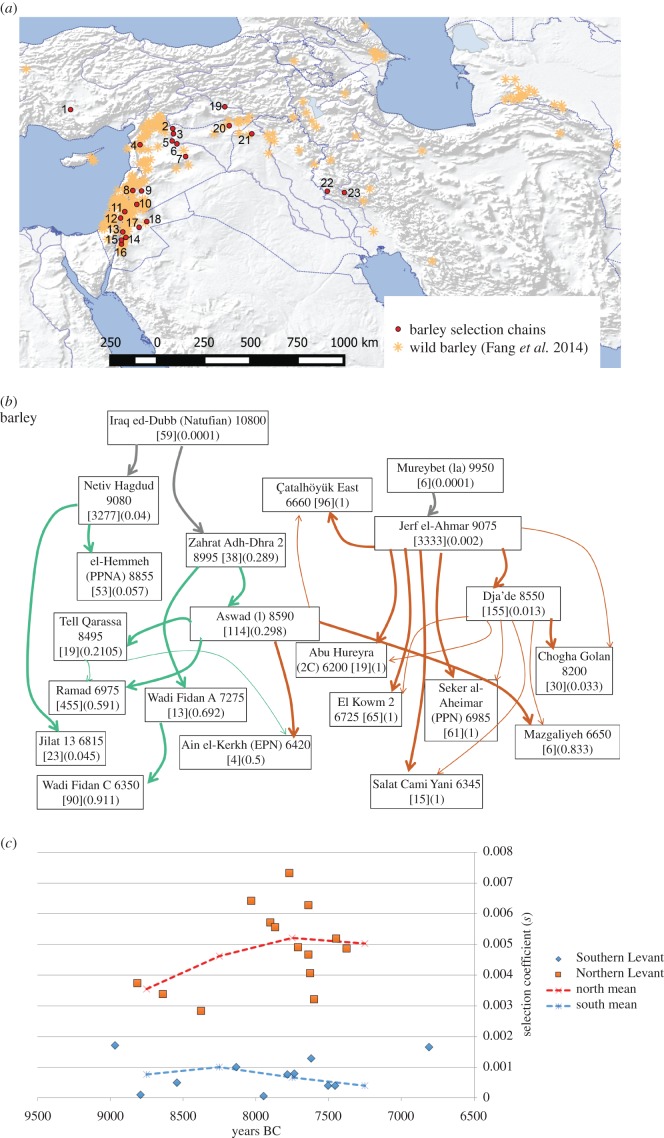
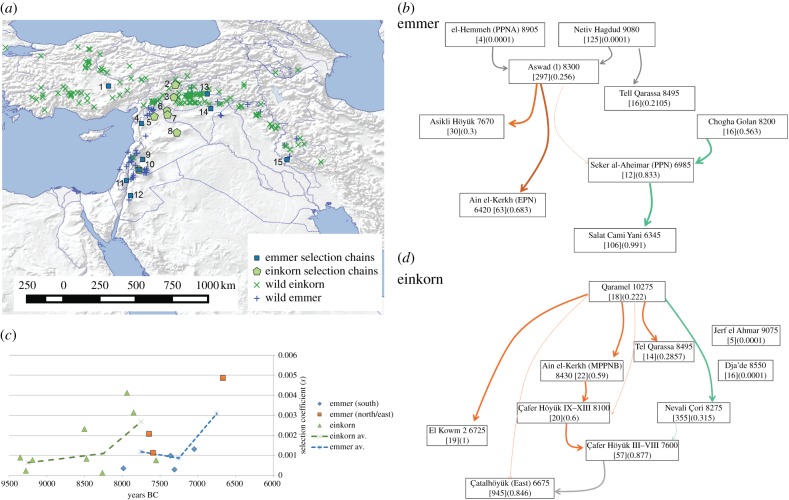
Domestication is the process by which plants or animals evolved to fit a human-managed environment, and it is marked by innovations in plant morphology and anatomy that are in turn correlated with new human behaviours and technologies for harvesting, storage and field preparation. Archaeobotanical evidence has revealed that domestication was a protracted process taking thousands of plant generations. Within this protracted process there were changes in the selection pressures for domestication traits as well as variation across a geographic mosaic of wild and cultivated populations. Quantitative data allow us to estimate the changing selection coefficients for the evolution of non-shattering (domestic-type seed dispersal) in Asian rice (Oryza sativa L.), barley (Hordeum vulgare L.), emmer wheat (Triticum dicoccon (Shrank) Schübl.) and einkorn wheat (Triticum monococcum L.). These data indicate that selection coefficients tended to be low, but also that there were inflection points at which selection increased considerably. For rice, selection coefficients of the order of 0.001 prior to 5500 BC shifted to greater than 0.003 between 5000 and 4500 BC, before falling again as the domestication process ended 4000-3500 BC. In barley and the two wheats selection was strongest between 8500 and 7500 BC. The slow start of domestication may indicate that initial selection began in the Pleistocene glacial era.This article is part of the themed issue 'Process and pattern in innovations from cells to societies'.
Keywords: China; Near East; archaeobotany; archaeology; niche construction; origins of agriculture.
© 2017 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
References
-
- Rindos D. 1984. The origins of agriculture. An evolutionary perspective. London, UK: Academic Press.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources