

Optogenetic Neuronal Silencing in Drosophila during Visual Processing
- PMID: 29061981
- PMCID: PMC5653863
- DOI: 10.1038/s41598-017-14076-7
Optogenetic Neuronal Silencing in Drosophila during Visual Processing
Abstract
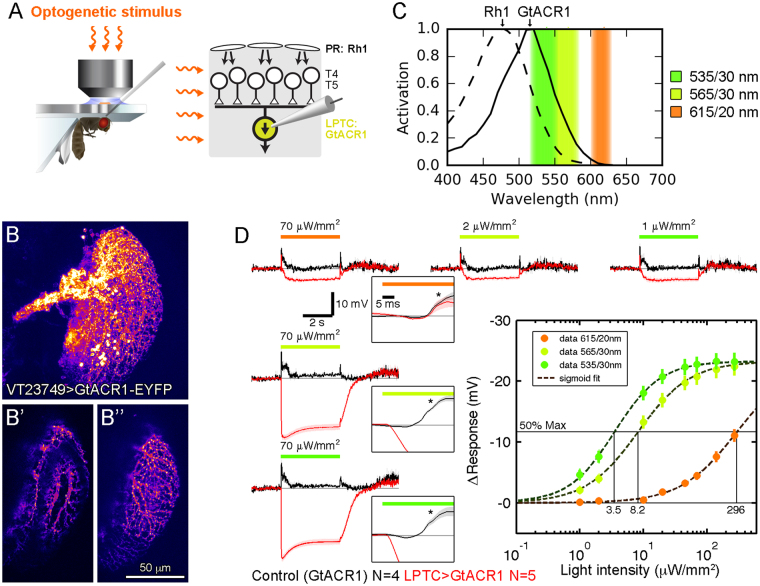
Optogenetic channels and ion pumps have become indispensable tools in neuroscience to manipulate neuronal activity and thus to establish synaptic connectivity and behavioral causality. Inhibitory channels are particularly advantageous to explore signal processing in neural circuits since they permit the functional removal of selected neurons on a trial-by-trial basis. However, applying these tools to study the visual system poses a considerable challenge because the illumination required for their activation usually also stimulates photoreceptors substantially, precluding the simultaneous probing of visual responses. Here, we explore the utility of the recently discovered anion channelrhodopsins GtACR1 and GtACR2 for application in the visual system of Drosophila. We first characterized their properties using a larval crawling assay. We further obtained whole-cell recordings from cells expressing GtACR1, which mediated strong and light-sensitive photocurrents. Finally, using physiological recordings and a behavioral readout, we demonstrate that GtACR1 enables the fast and reversible silencing of genetically targeted neurons within circuits engaged in visual processing.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
