

Monoaminergic Modulation of Motor Cortex Function
- PMID: 29062274
- PMCID: PMC5640772
- DOI: 10.3389/fncir.2017.00072
Monoaminergic Modulation of Motor Cortex Function
Abstract
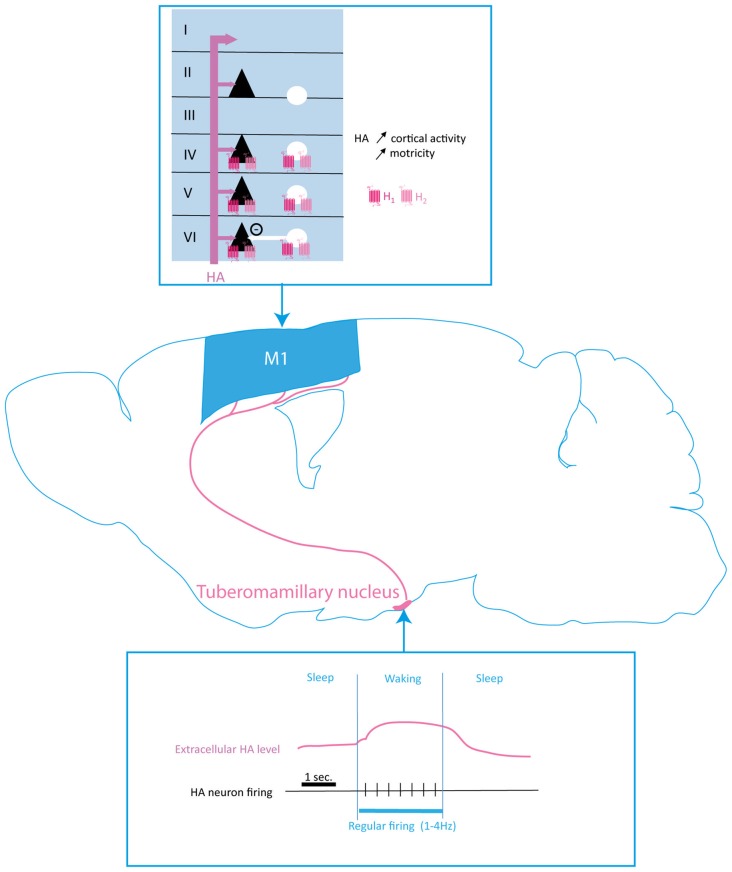
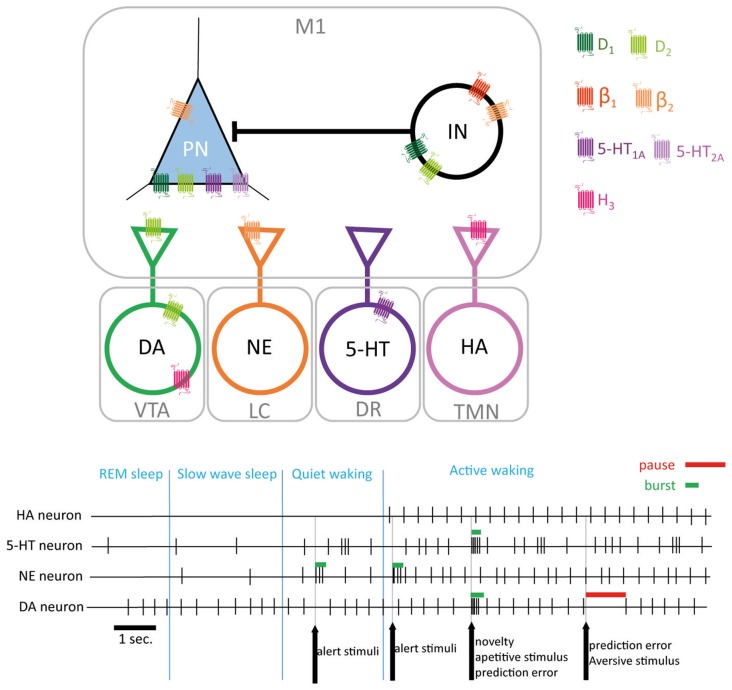
Elaboration of appropriate responses to behavioral situations rests on the ability of selecting appropriate motor outcomes in accordance to specific environmental inputs. To this end, the primary motor cortex (M1) is a key structure for the control of voluntary movements and motor skills learning. Subcortical loops regulate the activity of the motor cortex and thus contribute to the selection of appropriate motor plans. Monoamines are key mediators of arousal, attention and motivation. Their firing pattern enables a direct encoding of different states thus promoting or repressing the selection of actions adapted to the behavioral context. Monoaminergic modulation of motor systems has been extensively studied in subcortical circuits. Despite evidence of converging projections of multiple neurotransmitters systems in the motor cortex pointing to a direct modulation of local circuits, their contribution to the execution and learning of motor skills is still poorly understood. Monoaminergic dysregulation leads to impaired plasticity and motor function in several neurological and psychiatric conditions, thus it is critical to better understand how monoamines modulate neural activity in the motor cortex. This review aims to provide an update of our current understanding on the monoaminergic modulation of the motor cortex with an emphasis on motor skill learning and execution under physiological conditions.
Keywords: dopamine; histamine; monoamines; motor cortex; norepinephrine; serotonin.
Figures
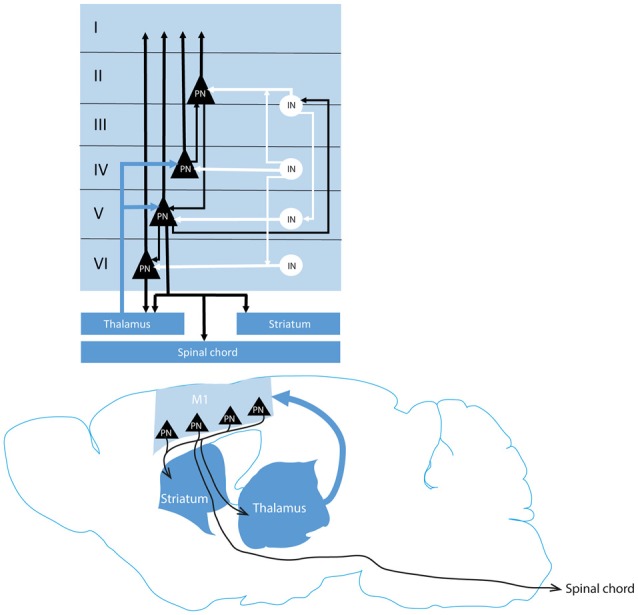
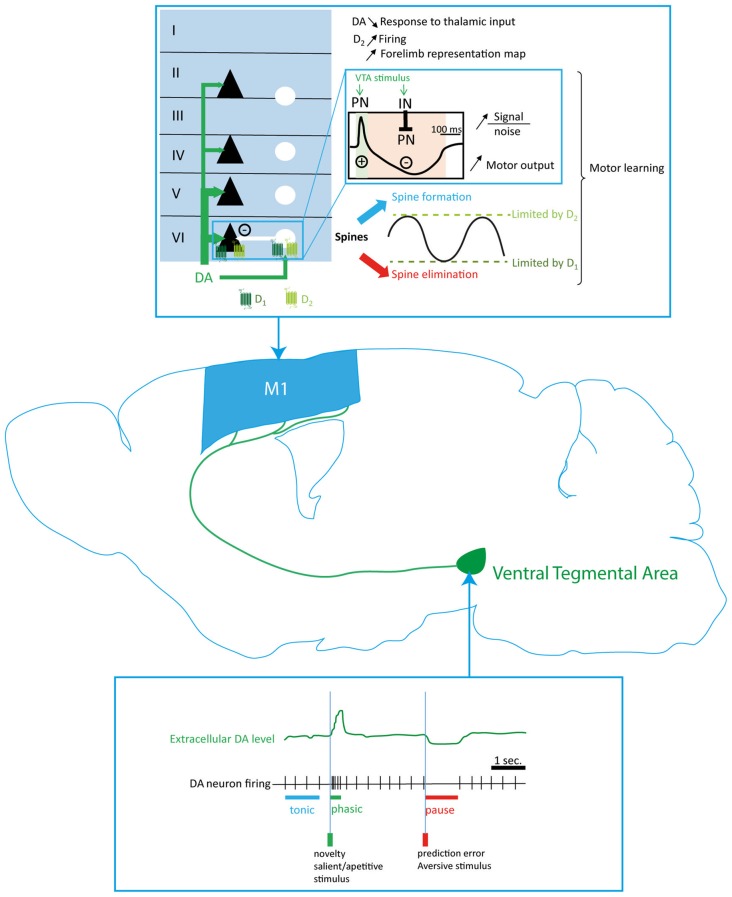
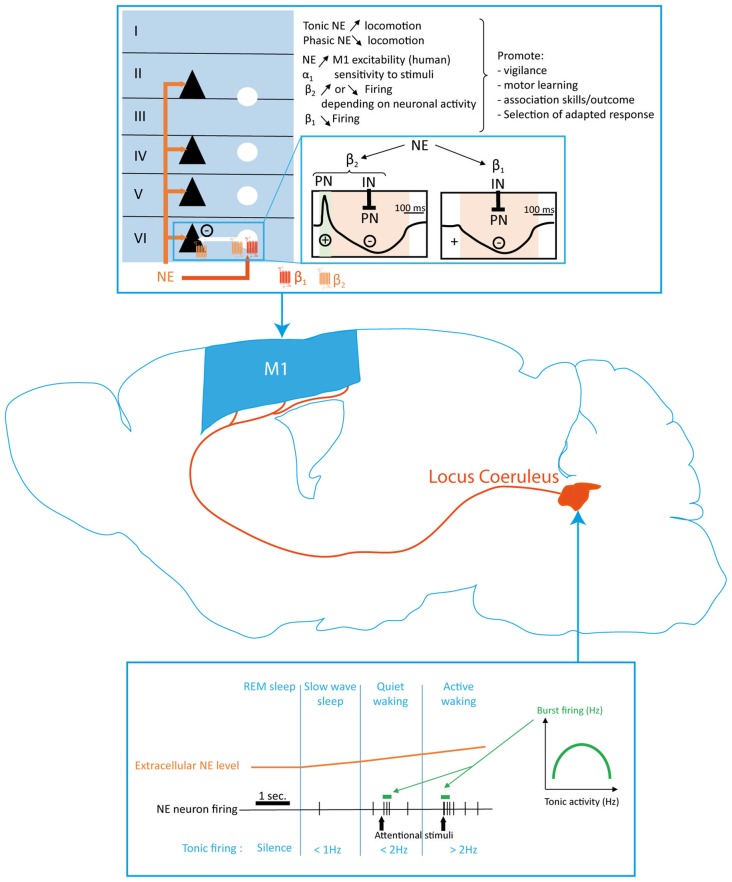
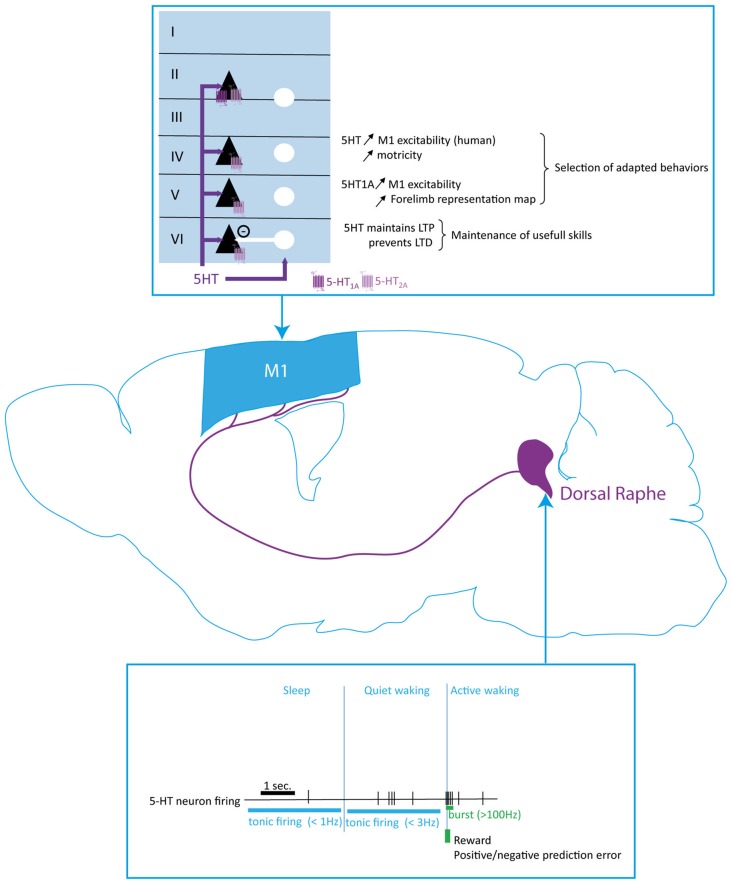
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources