

Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome
- PMID: 29066723
- PMCID: PMC5655019
- DOI: 10.1038/s41598-017-13832-z
Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome
Abstract
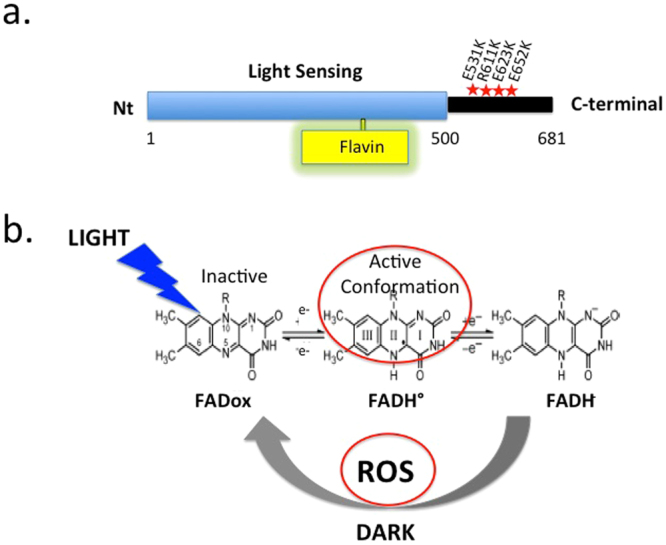
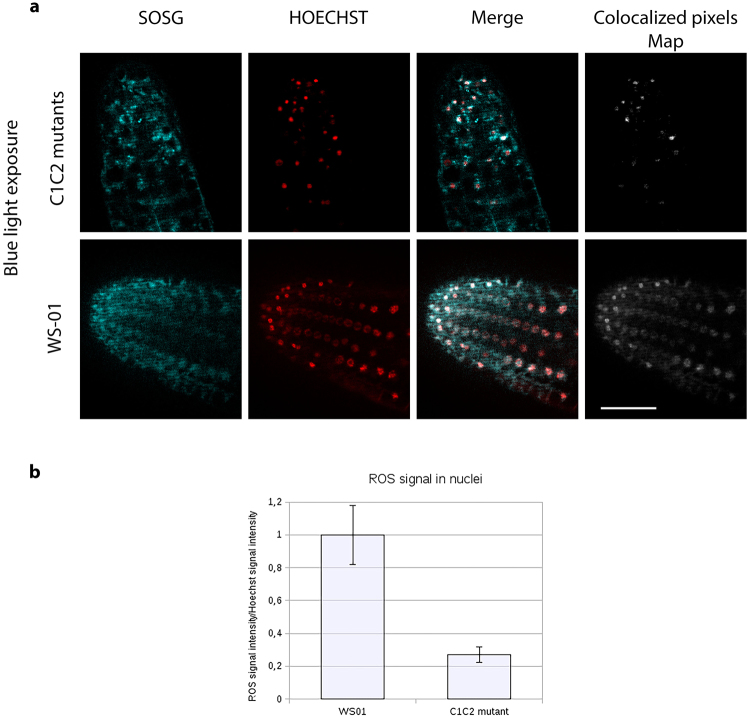
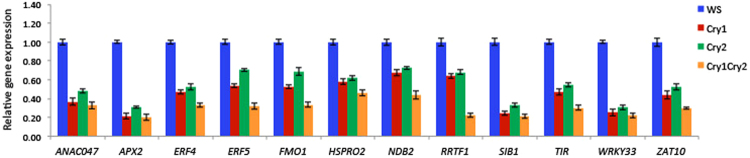
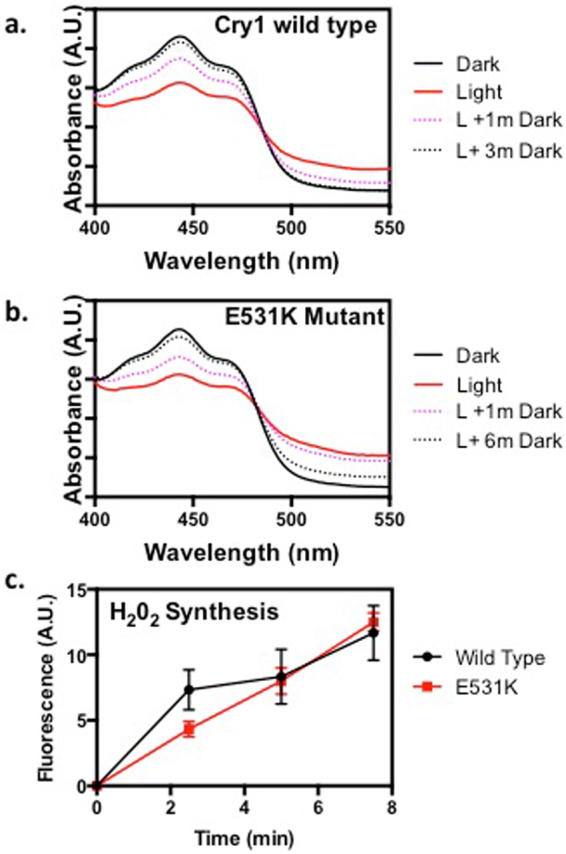
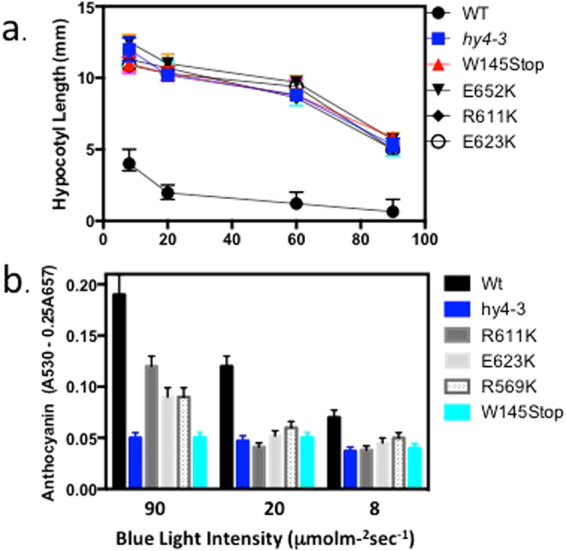
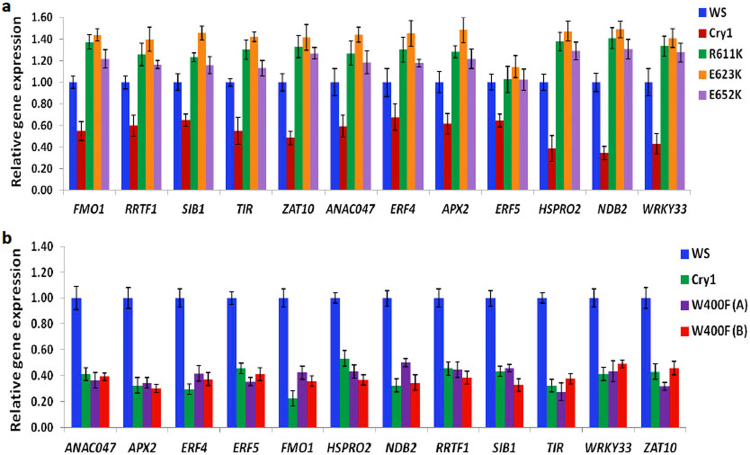
Cryptochromes are evolutionarily conserved blue light receptors with many roles throughout plant growth and development. They undergo conformational changes in response to light enabling interaction with multiple downstream signaling partners. Recently, it has been shown that cryptochromes also synthesize reactive oxygen species (ROS) in response to light, suggesting the possibility of an alternate signaling mechanism. Here we show by fluorescence imaging and microscopy that H202 and ROS accumulate in the plant nucleus after cryptochrome activation. They induce ROS-regulated transcripts including for genes implicated in pathogen defense, biotic and abiotic stress. Mutant cryptochrome alleles that are non-functional in photomorphogenesis retain the capacity to induce ROS-responsive phenotypes. We conclude that nuclear biosynthesis of ROS by cryptochromes represents a new signaling paradigm that complements currently known mechanisms. This may lead to novel applications using blue light induced oxidative bursts to prime crop plants against the deleterious effects of environmental stresses and toxins.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
