

The yeast form of the fungus Candida albicans promotes persistence in the gut of gnotobiotic mice
- PMID: 29069103
- PMCID: PMC5673237
- DOI: 10.1371/journal.ppat.1006699
The yeast form of the fungus Candida albicans promotes persistence in the gut of gnotobiotic mice
Abstract
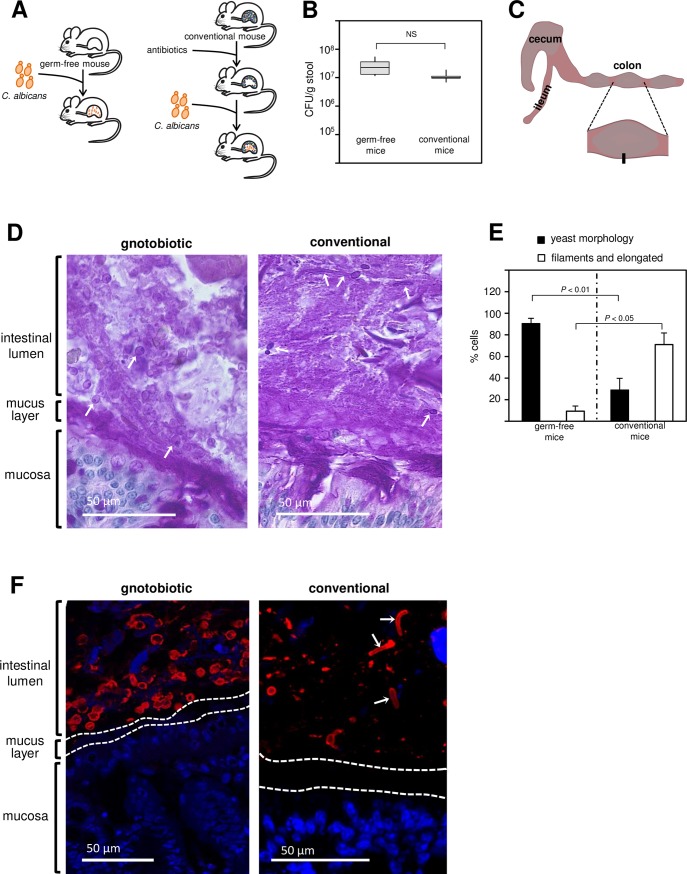
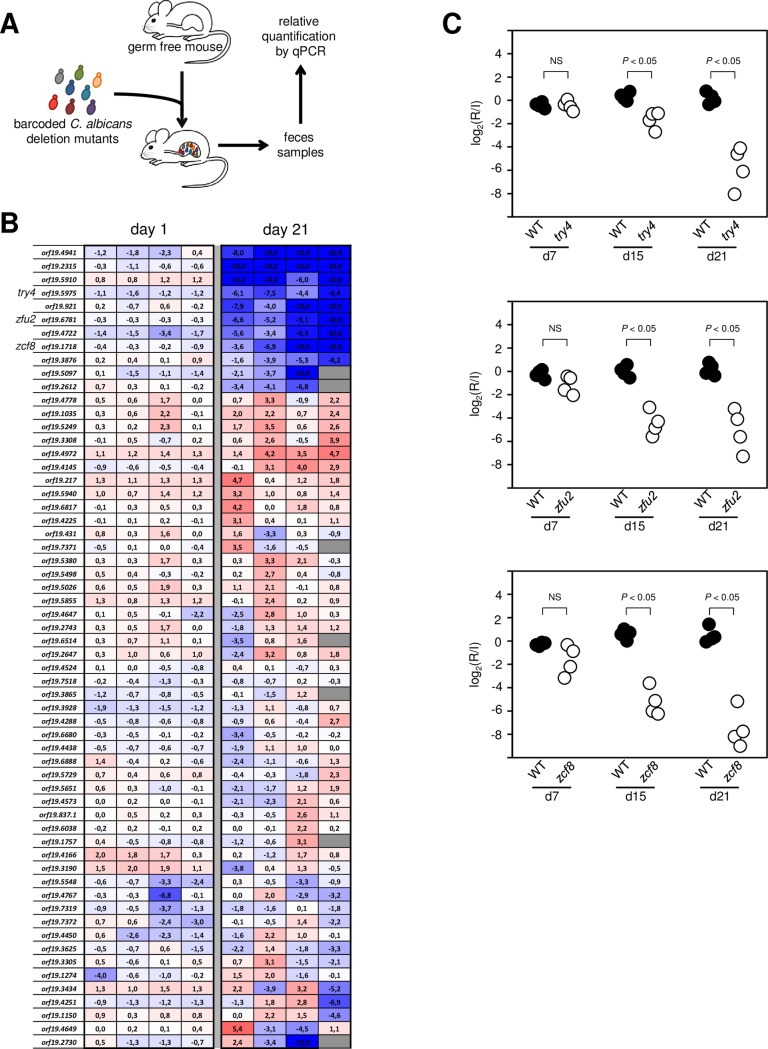
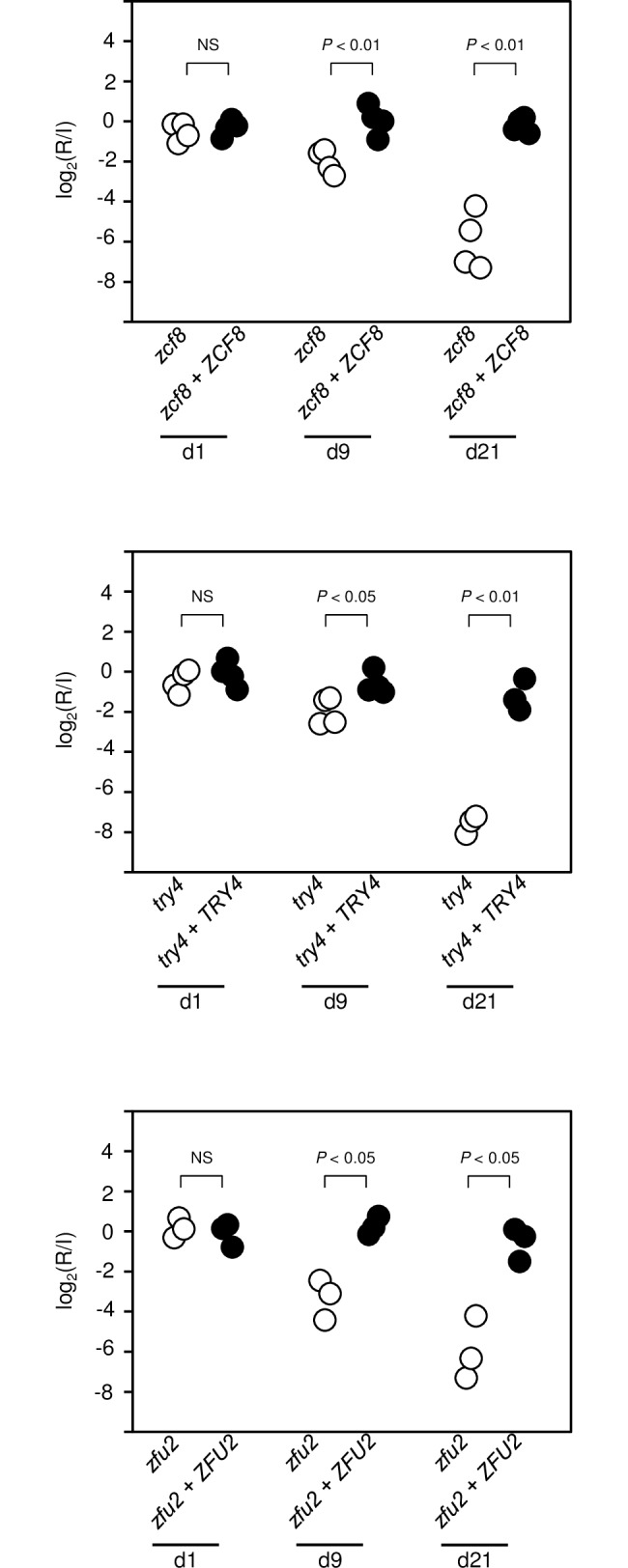
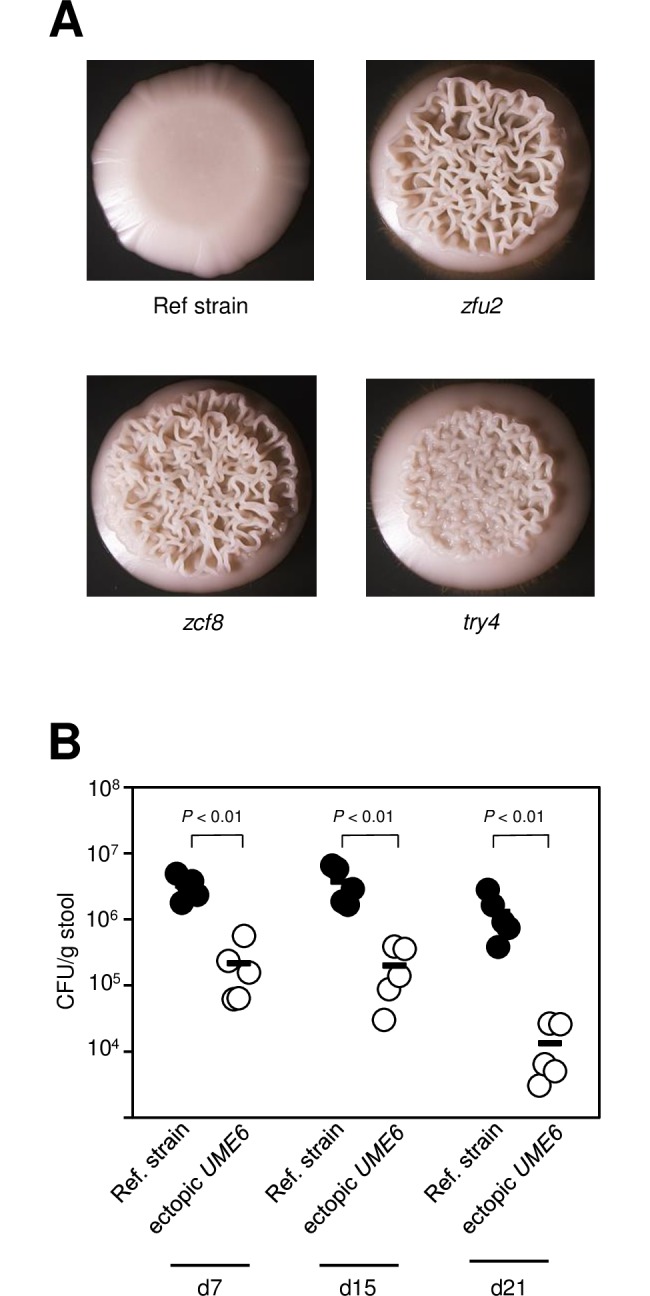
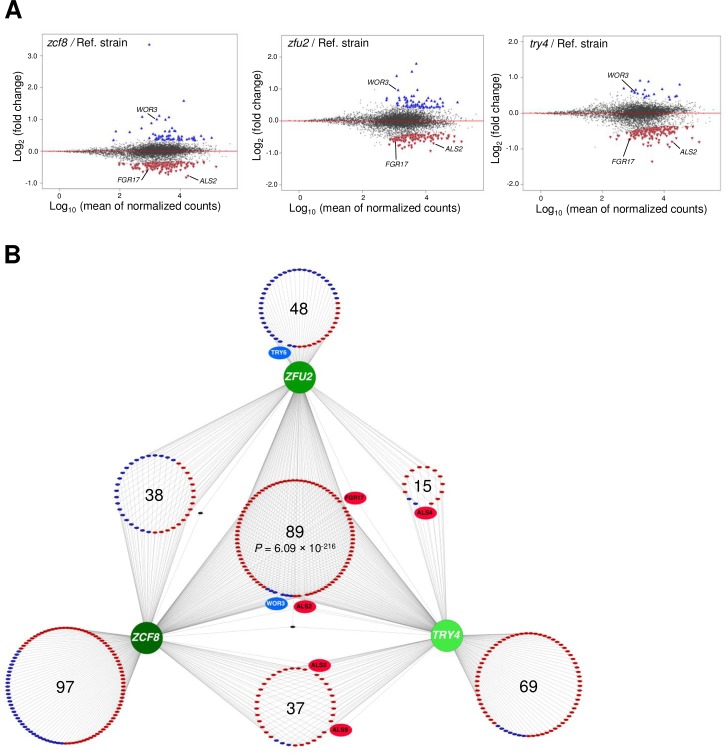
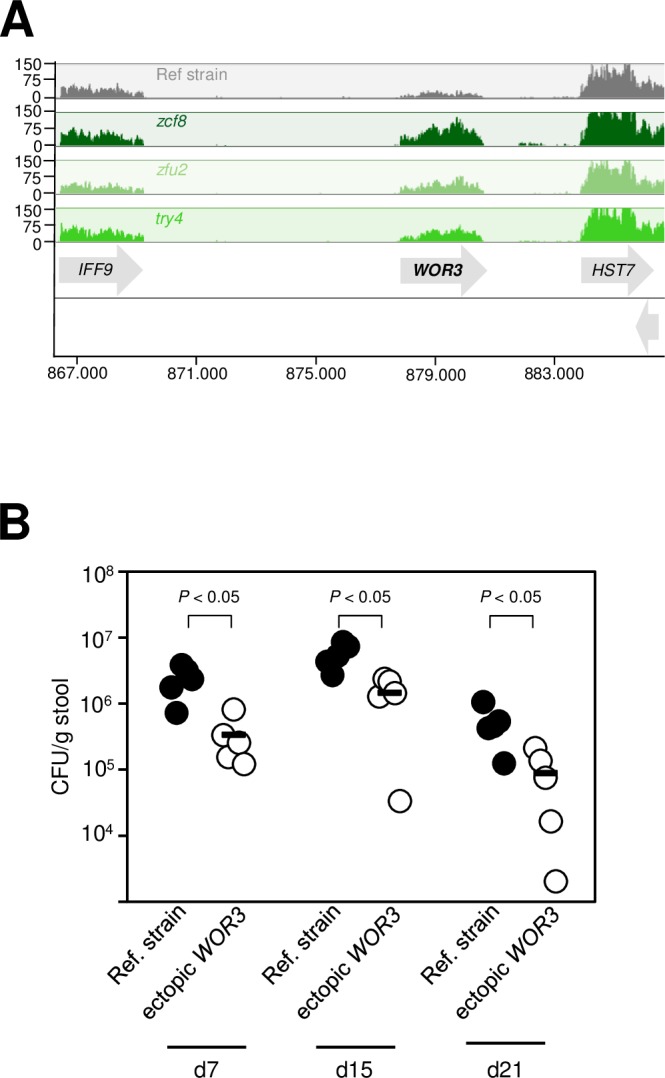
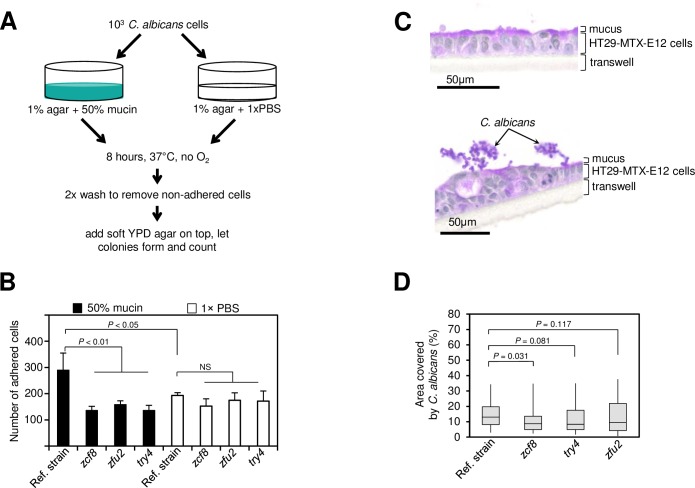
Many microorganisms that cause systemic, life-threatening infections in humans reside as harmless commensals in our digestive tract. Yet little is known about the biology of these microbes in the gut. Here, we visualize the interface between the human commensal and pathogenic fungus Candida albicans and the intestine of mice, a surrogate host. Because the indigenous mouse microbiota restricts C. albicans settlement, we compared the patterns of colonization in the gut of germ free and antibiotic-treated conventionally raised mice. In contrast to the heterogeneous morphologies found in the latter, we establish that in germ free animals the fungus almost uniformly adopts the yeast cell form, a proxy of its commensal state. By screening a collection of C. albicans transcription regulator deletion mutants in gnotobiotic mice, we identify several genes previously unknown to contribute to in vivo fitness. We investigate three of these regulators-ZCF8, ZFU2 and TRY4-and show that indeed they favor the yeast form over other morphologies. Consistent with this finding, we demonstrate that genetically inducing non-yeast cell morphologies is detrimental to the fitness of C. albicans in the gut. Furthermore, the identified regulators promote adherence of the fungus to a surface covered with mucin and to mucus-producing intestinal epithelial cells. In agreement with this result, histology sections indicate that C. albicans dwells in the murine gut in close proximity to the mucus layer. Thus, our findings reveal a set of regulators that endows C. albicans with the ability to endure in the intestine through multiple mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Chudnovskiy A, Mortha A, Kana V, Kennard A, Ramirez JD, et al. (2016) Host-Protozoan Interactions Protect from Mucosal Infections through Activation of the Inflammasome. Cell 167: 444–456 e414. doi: 10.1016/j.cell.2016.08.076 - DOI - PMC - PubMed
-
- Lukes J, Stensvold CR, Jirku-Pomajbikova K, Wegener Parfrey L (2015) Are Human Intestinal Eukaryotes Beneficial or Commensals? PLoS Pathog 11: e1005039 doi: 10.1371/journal.ppat.1005039 - DOI - PMC - PubMed
-
- Underhill DM, Iliev ID (2014) The mycobiota: interactions between commensal fungi and the host immune system. Nat Rev Immunol 14: 405–416. doi: 10.1038/nri3684 - DOI - PMC - PubMed
-
- Wheeler ML, Limon JJ, Bar AS, Leal CA, Gargus M, et al. (2016) Immunological Consequences of Intestinal Fungal Dysbiosis. Cell Host Microbe 19: 865–873. doi: 10.1016/j.chom.2016.05.003 - DOI - PMC - PubMed
-
- Kumamoto CA (2016) The Fungal Mycobiota: Small Numbers, Large Impacts. Cell Host Microbe 19: 750–751. doi: 10.1016/j.chom.2016.05.018 - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
