

Lateralized Feeding Behavior is Associated with Asymmetrical Neuroanatomy and Lateralized Gene Expressions in the Brain in Scale-Eating Cichlid Fish
- PMID: 29069363
- PMCID: PMC5737854
- DOI: 10.1093/gbe/evx218
Lateralized Feeding Behavior is Associated with Asymmetrical Neuroanatomy and Lateralized Gene Expressions in the Brain in Scale-Eating Cichlid Fish
Abstract
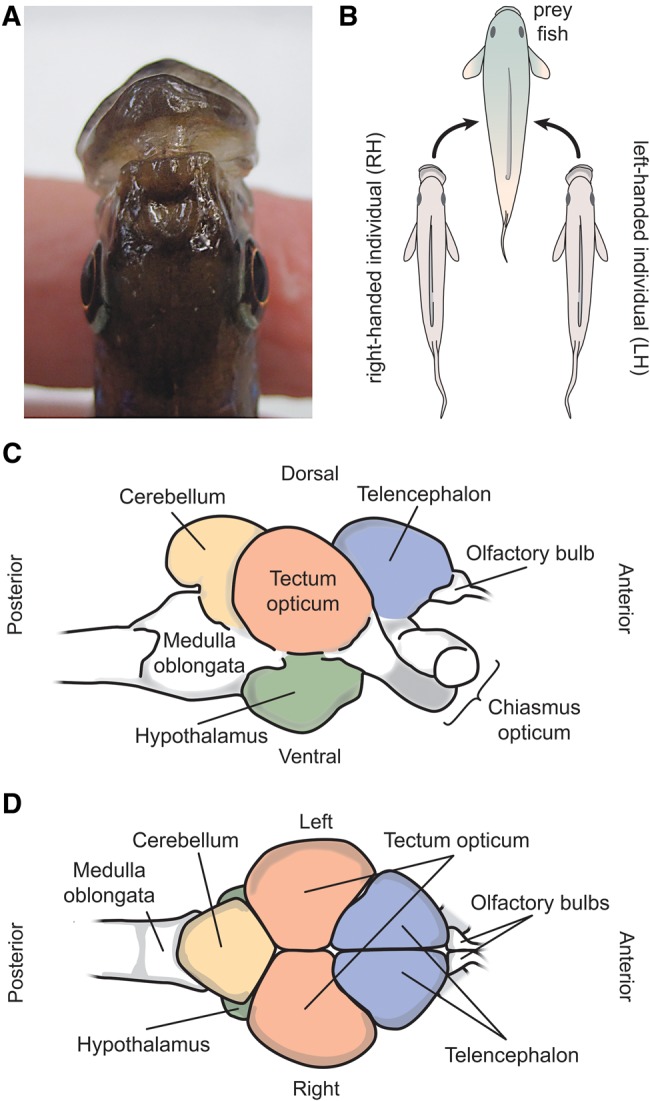
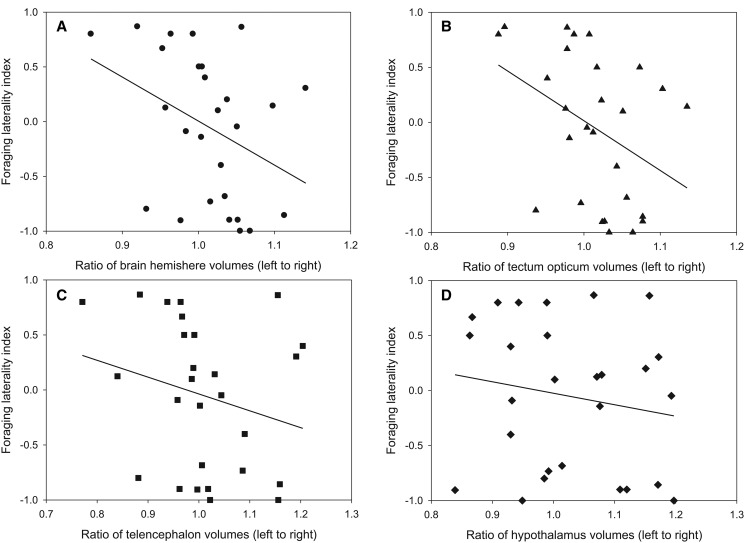
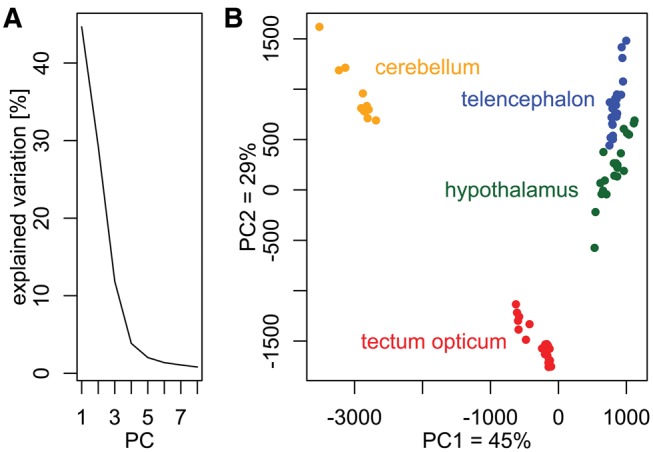
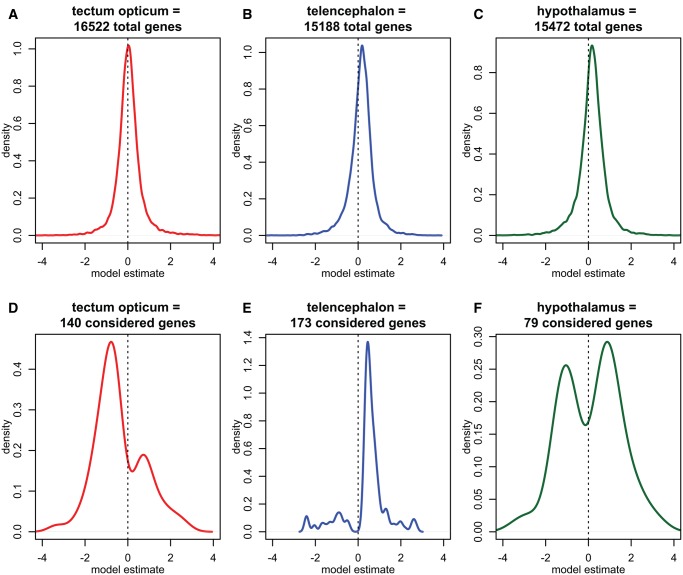
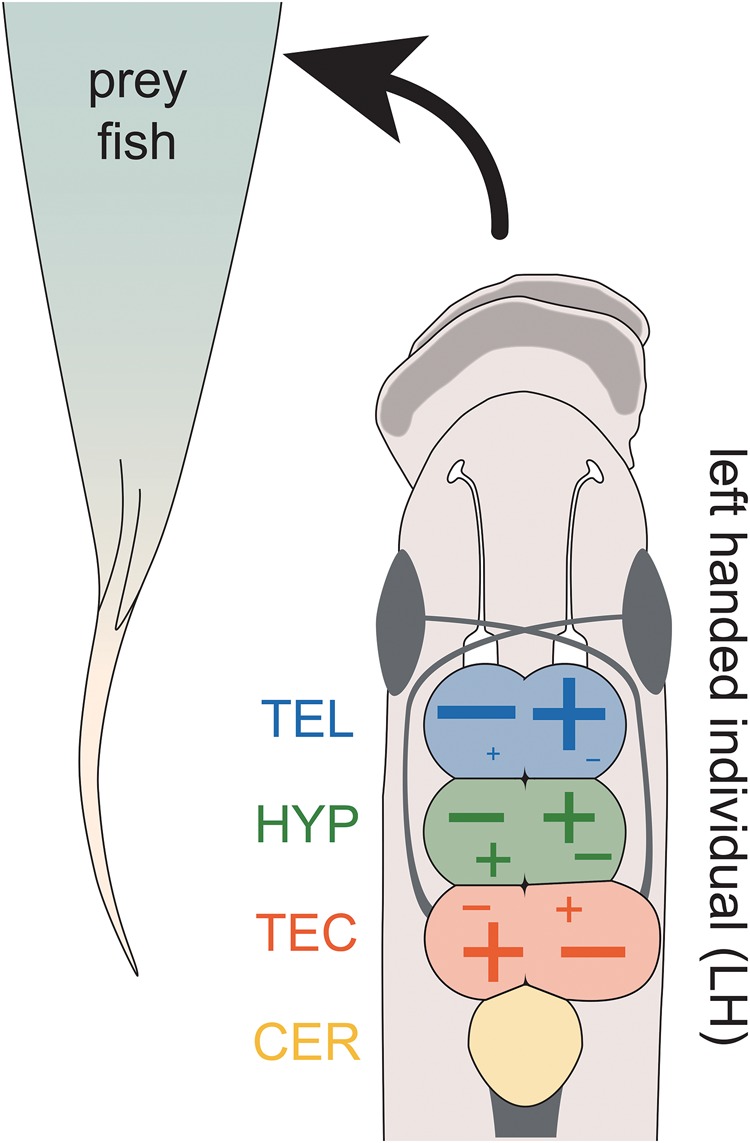
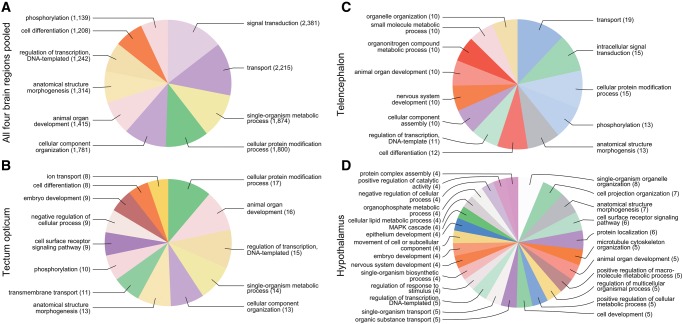
Lateralized behavior ("handedness") is unusual, but consistently found across diverse animal lineages, including humans. It is thought to reflect brain anatomical and/or functional asymmetries, but its neuro-molecular mechanisms remain largely unknown. Lake Tanganyika scale-eating cichlid fish, Perissodus microlepis show pronounced asymmetry in their jaw morphology as well as handedness in feeding behavior-biting scales preferentially only from one or the other side of their victims. This makes them an ideal model in which to investigate potential laterality in neuroanatomy and transcription in the brain in relation to behavioral handedness. After determining behavioral handedness in P. microlepis (preferred attack side), we estimated the volume of the hemispheres of brain regions and captured their gene expression profiles. Our analyses revealed that the degree of behavioral handedness is mirrored at the level of neuroanatomical asymmetry, particularly in the tectum opticum. Transcriptome analyses showed that different brain regions (tectum opticum, telencephalon, hypothalamus, and cerebellum) display distinct expression patterns, potentially reflecting their developmental interrelationships. For numerous genes in each brain region, their extent of expression differences between hemispheres was found to be correlated with the degree of behavioral lateralization. Interestingly, the tectum opticum and telencephalon showed divergent biases on the direction of up- or down-regulation of the laterality candidate genes (e.g., grm2) in the hemispheres, highlighting the connection of handedness with gene expression profiles and the different roles of these brain regions. Hence, handedness in predation behavior may be caused by asymmetric size of brain hemispheres and also by lateralized gene expressions in the brain.
Keywords: Perissodus microlepis; behavioral genetics/genomics; left-right asymmetry; neural structures; tectum opticum; telencephalon.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Ades C, Ramires EN.. 2002. Asymmetry of leg use during prey handling in the spider Scytodes globula (Scytodidae). J Insect Behav. 15:563–570.http://dx.doi.org/10.1023/A:1016337418472 - DOI
-
- Al-Akel AS, Guthrie DM, Banks JR.. 1986. Motor responses to localized electrical stimulation of the tectum in the freshwater perch (Perca fluviatilis). Neuroscience 19(4):1381–1391.http://dx.doi.org/10.1016/0306-4522(86)90150-8 - DOI - PubMed
-
- Alcock J. 2009. Animal behavior. Sunderland: Sinauer Associates.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
