

A peptide antagonist of Prep1-p160 interaction improves ceramide-induced insulin resistance in skeletal muscle cells
- PMID: 29069751
- PMCID: PMC5641094
- DOI: 10.18632/oncotarget.18286
A peptide antagonist of Prep1-p160 interaction improves ceramide-induced insulin resistance in skeletal muscle cells
Abstract
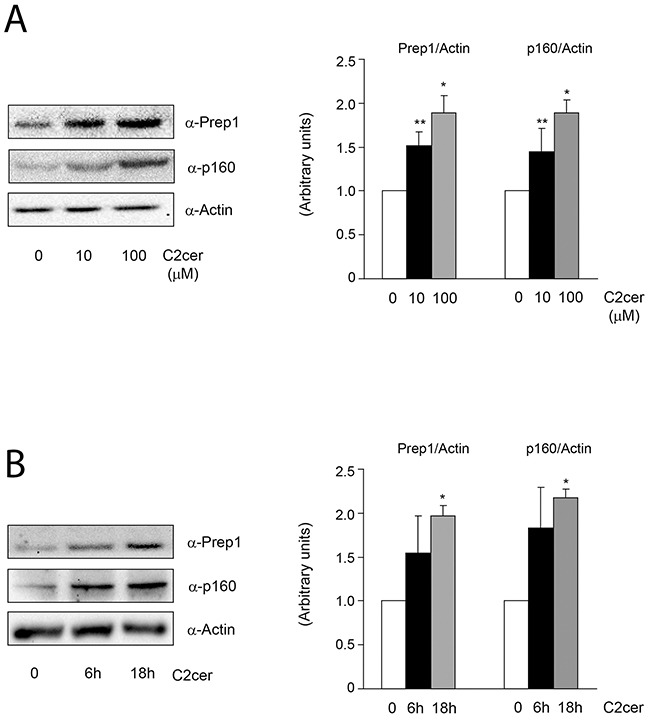
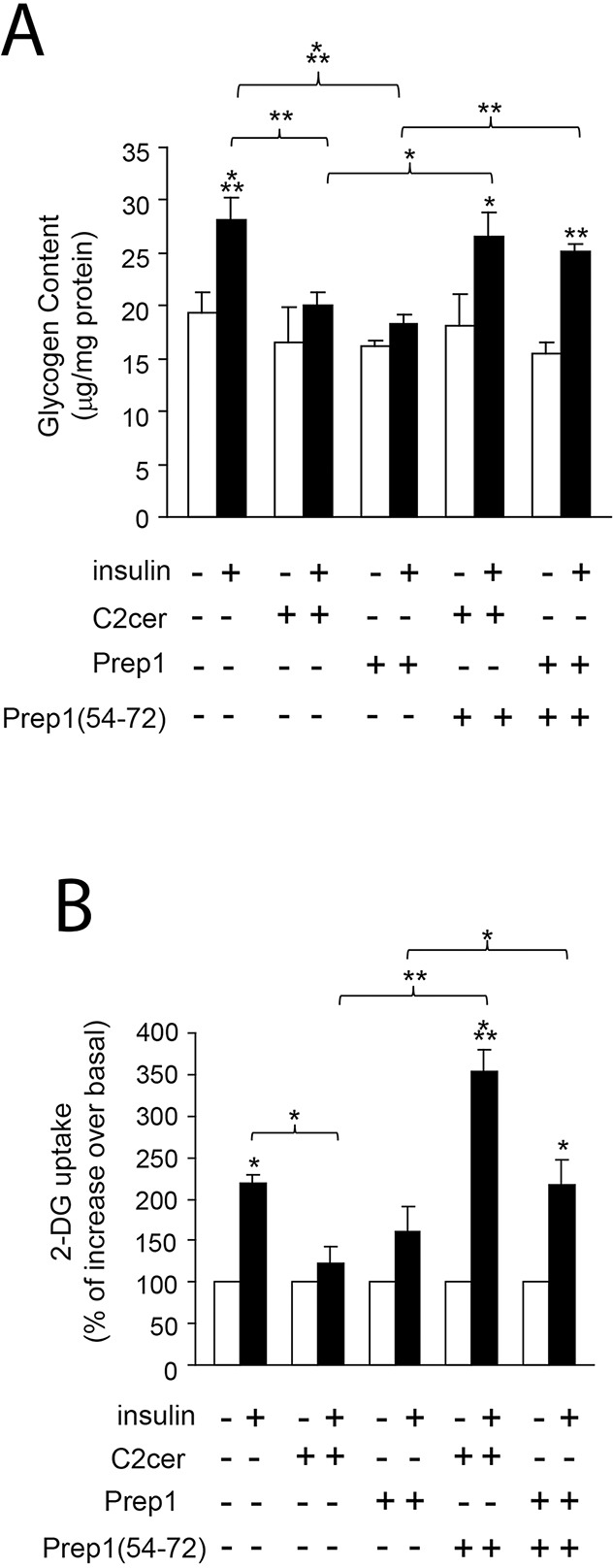
Prep1 is a homeodomain transcription factor belonging to the TALE protein family. Its overexpression affects glucose metabolism in several tissues. In particular, in skeletal muscle tissue the interaction of Prep1 with its cofactor p160 impairs GLUT4 expression and glucose uptake. In this study, we show that ceramides (C2cer), a class of lipids antagonizing insulin signalling, increase the levels of Prep1 and p160 in a dose and time-dependent fashion in L6 cells and induce their association by 80%. We find that C2cer exposure inhibits insulin receptor, IRS1 and Akt phosphorylation and reduces insulin-stimulated glycogen content and glucose uptake by 1.3- and 2.1-fold, respectively. The synthetic Prep1(54-72) peptide, mimicking the Prep1 region involved in the interaction with p160, reduces in vitro Prep1-p160 binding in a dose-dependent way (IC50 = 0.20μM). In C2cer-treated L6 cells, 10μM Prep1(54-72) restores insulin signalling impaired by ceramide treatment. Prep1 overexpressing L6 cells display similar metabolic alterations observed in ceramide-treated L6 cells and the presence of Prep1(54-72) mitigates these events. All these findings suggest that disruption of the Prep1/p160 molecular interaction enhances insulin sensitivity impaired by ceramides in skeletal muscle cells and indicate this complex as an important target for type 2 diabetes.
Keywords: Prep1; Prep1(54-72) peptide; ceramide; insulin signalling; p160.
Conflict of interest statement
CONFLICTS OF INTEREST All authors have no conflicts of interest.
Figures





References
-
- Summers SA. Ceramides in insulin resistance and lipotoxicity. Prog Lipid Res. 2006;45:42–72. - PubMed
-
- Kanety H, Hemi R, Papa MZ, Karasik A. Sphingomyelinase and ceramide suppress insulin-induced tyrosine phosphorylation of the insulin receptor substrate-1. The Journal of biological chemistry. 1996;271:9895–9897. - PubMed
-
- Hajduch E, Balendran A, Batty IH, Litherland GJ, Blair AS, Downes CP, Hundal HS. Ceramide impairs the insulin-dependent membrane recruitment of protein kinase B leading to a loss in downstream signalling in L6 skeletal muscle cells. Diabetologia. 2001;44:173–183. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
