

Plant hormone transporters: what we know and what we would like to know
- PMID: 29070024
- PMCID: PMC5655956
- DOI: 10.1186/s12915-017-0443-x
Plant hormone transporters: what we know and what we would like to know
Abstract
Hormone transporters are crucial for plant hormone action, which is underlined by severe developmental and physiological impacts caused by their loss-of-function mutations. Here, we summarize recent knowledge on the individual roles of plant hormone transporters in local and long-distance transport. Our inventory reveals that many hormones are transported by members of distinct transporter classes, with an apparent dominance of the ATP-binding cassette (ABC) family and of the Nitrate transport1/Peptide transporter family (NPF). The current need to explore further hormone transporter regulation, their functional interaction, transport directionalities, and substrate specificities is briefly reviewed.
Conflict of interest statement
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
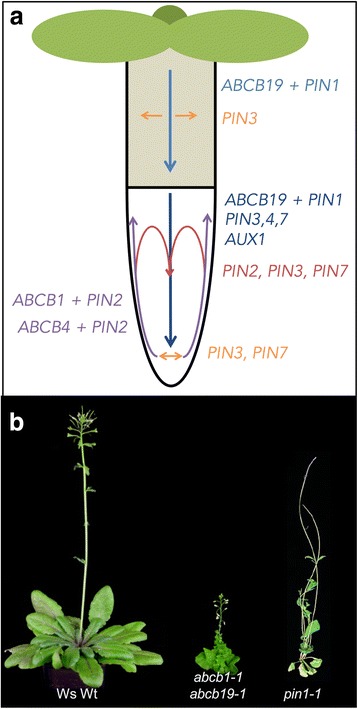
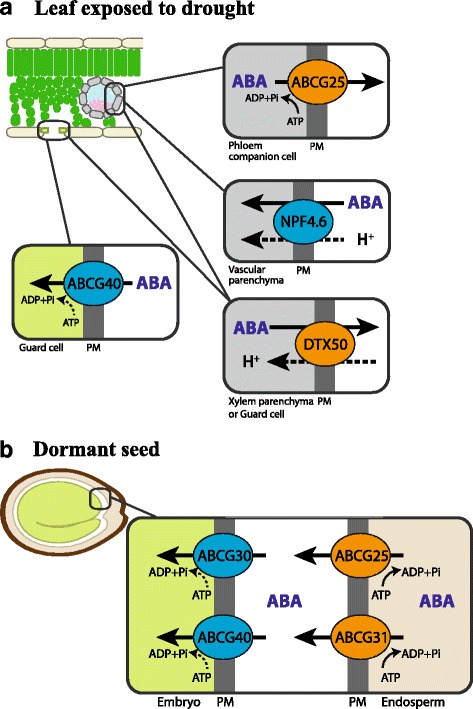


References
-
- Darwin C, Darwin F. The power of movements in plants. London: Murray; 1880.
-
- Christie JM, Murphy AS. Shoot phototropism in higher plants: new light through old concepts. Am J Bot. 2013;100:35–46. - PubMed
-
- Bauer H, Ache P, Lautner S, Fromm J, Hartung W, Al-Rasheid KA, et al. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr Biol. 2013;23:53–7. - PubMed
-
- Davies PJ. The plant hormones: their nature, occurrence, and functions. In: Davies PJ, editor. The plant hormones: biosynthesis, signal transduction, action! 3. Dordrecht: Springer Netherlands; 2010. pp. 1–15.
-
- Yang SF, Hoffmann NE. Ethylene biosynthesis and its regulation in higher plants. Annu Rev Plant Physiol. 1984;35:155–89.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
