

Minimal and RNA-free RNase P in Aquifex aeolicus
- PMID: 29073018
- PMCID: PMC5651759
- DOI: 10.1073/pnas.1707862114
Minimal and RNA-free RNase P in Aquifex aeolicus
Abstract
RNase P is an essential tRNA-processing enzyme in all domains of life. We identified an unknown type of protein-only RNase P in the hyperthermophilic bacterium Aquifex aeolicus: Without an RNA subunit and the smallest of its kind, the 23-kDa polypeptide comprises a metallonuclease domain only. The protein has RNase P activity in vitro and rescued the growth of Escherichia coli and Saccharomyces cerevisiae strains with inactivations of their more complex and larger endogenous ribonucleoprotein RNase P. Homologs of Aquifex RNase P (HARP) were identified in many Archaea and some Bacteria, of which all Archaea and most Bacteria also encode an RNA-based RNase P; activity of both RNase P forms from the same bacterium or archaeon could be verified in two selected cases. Bioinformatic analyses suggest that A. aeolicus and related Aquificaceae likely acquired HARP by horizontal gene transfer from an archaeon.
Keywords: Aquifex aeolicus; HARP; protein-only RNase P; tRNA processing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
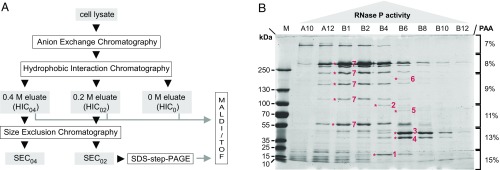
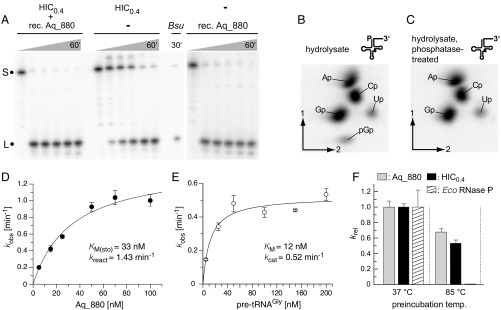
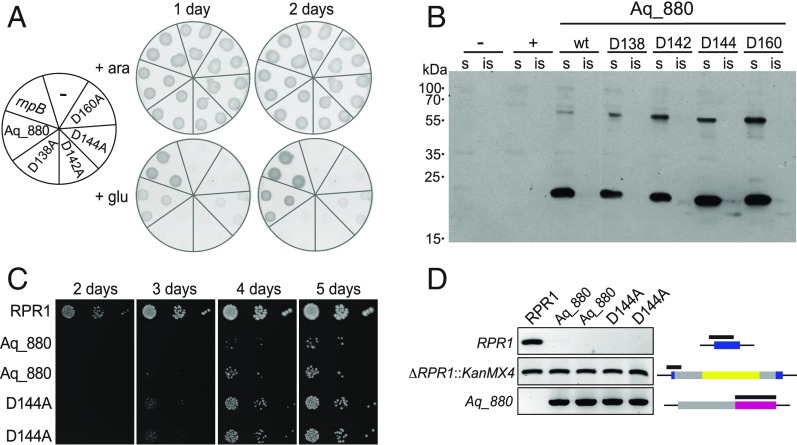
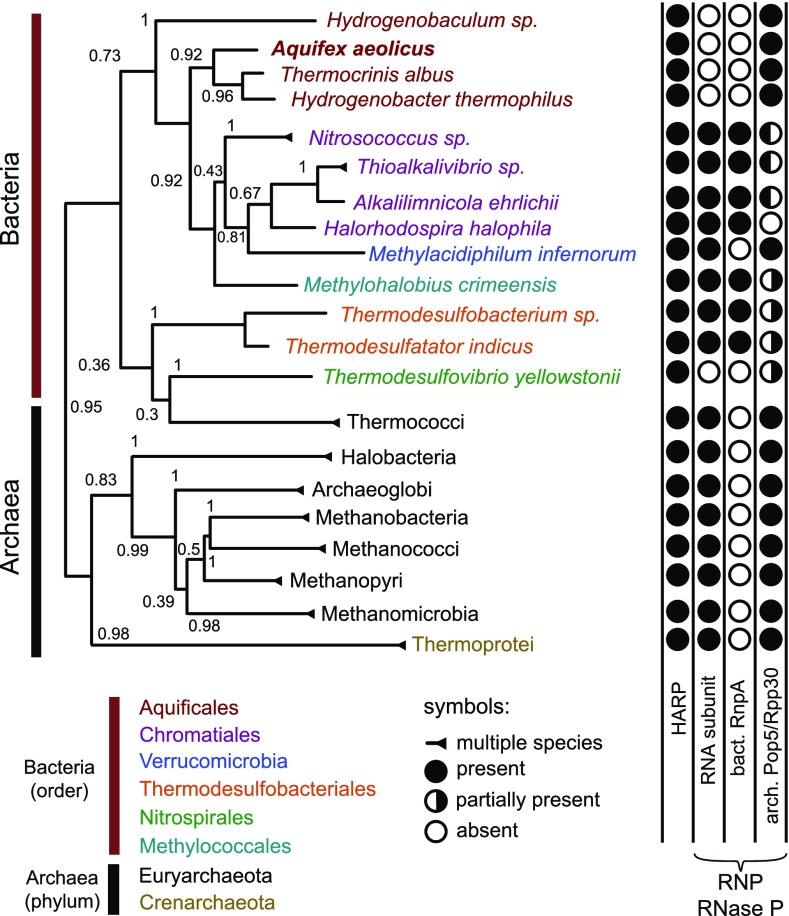
References
-
- Hartmann E, Hartmann RK. The enigma of ribonuclease P evolution. Trends Genet. 2003;19:561–569. - PubMed
-
- Holzmann J, et al. RNase P without RNA: Identification and functional reconstitution of the human mitochondrial tRNA processing enzyme. Cell. 2008;135:462–474. - PubMed
-
- Gobert A, et al. A single Arabidopsis organellar protein has RNase P activity. Nat Struct Mol Biol. 2010;17:740–744. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
