

Identification of novel substrates of Shigella T3SA through analysis of its virulence plasmid-encoded secretome
- PMID: 29073283
- PMCID: PMC5658099
- DOI: 10.1371/journal.pone.0186920
Identification of novel substrates of Shigella T3SA through analysis of its virulence plasmid-encoded secretome
Abstract
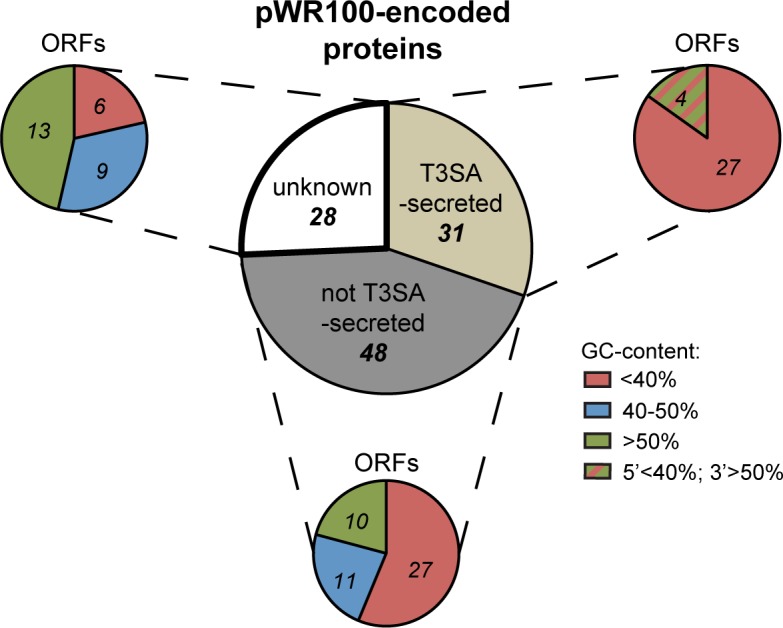
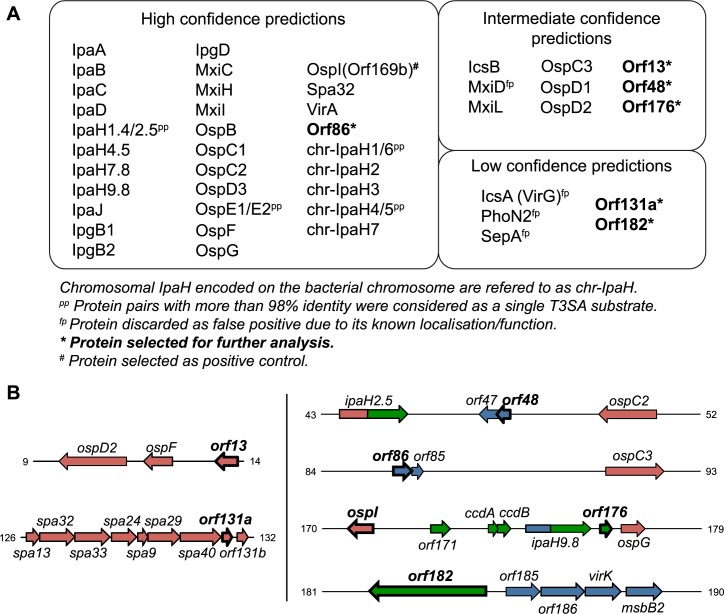
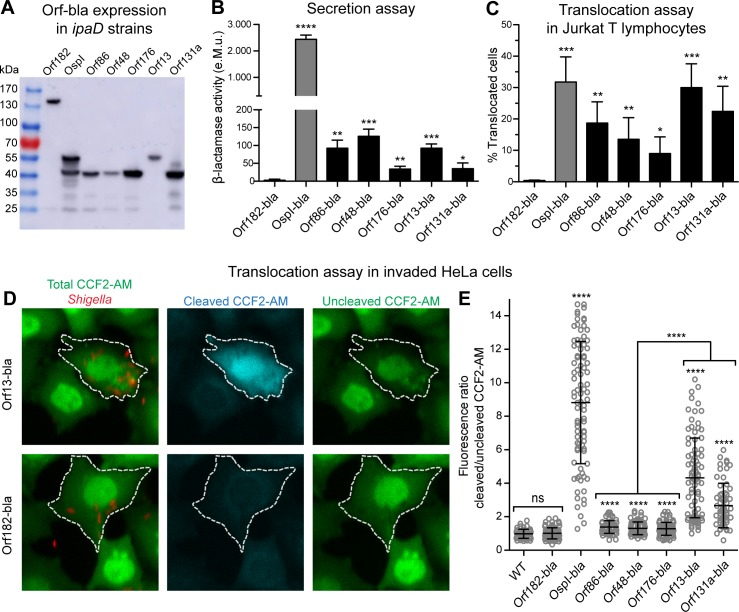
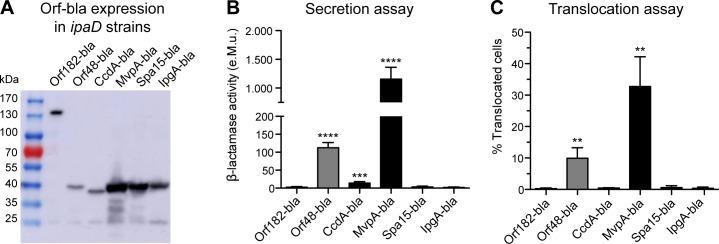
Many human Gram-negative bacterial pathogens express a Type Three Secretion Apparatus (T3SA), including among the most notorious Shigella spp., Salmonella enterica, Yersinia enterocolitica and enteropathogenic Escherichia coli (EPEC). These bacteria express on their surface multiple copies of the T3SA that mediate the delivery into host cells of specific protein substrates critical to pathogenesis. Shigella spp. are Gram-negative bacterial pathogens responsible for human bacillary dysentery. The effector function of several Shigella T3SA substrates has largely been studied but their potential cellular targets are far from having been comprehensively delineated. In addition, it is likely that some T3SA substrates have escaped scrutiny as yet. Indeed, sequencing of the virulence plasmid of Shigella flexneri has revealed numerous open reading frames with unknown functions that could encode additional T3SA substrates. Taking advantage of label-free mass spectrometry detection of proteins secreted by a constitutively secreting strain of S. flexneri, we identified five novel substrates of the T3SA. We further confirmed their secretion through the T3SA and translocation into host cells using β-lactamase assays. The coding sequences of two of these novel T3SA substrates (Orf13 and Orf131a) have a guanine-cytosine content comparable to those of T3SA components and effectors. The three other T3SA substrates identified (Orf48, Orf86 and Orf176) have significant homology with antitoxin moieties of type II Toxin-Antitoxin systems usually implicated in the maintenance of low copy plasmids. While Orf13 and Orf131a might constitute new virulence effectors contributing to S. flexneri pathogenicity, potential roles for the translocation into host cells of antitoxins or antitoxin-like proteins during Shigella infection are discussed.
Conflict of interest statement
Figures
References
-
- Galán JE, Lara-Tejero M, Marlovits TC, Wagner S. Bacterial type III secretion systems: specialized nanomachines for protein delivery into target cells. Annu Rev Microbiol. 2014;68: 415–438. doi: 10.1146/annurev-micro-092412-155725 - DOI - PMC - PubMed
-
- Ehsani S, Rodrigues CD, Enninga J. Turning on the spotlight—using light to monitor and characterize bacterial effector secretion and translocation. Curr Opin Microbiol. 2009;12: 24–30. doi: 10.1016/j.mib.2008.11.007 - DOI - PubMed
-
- Campbell-Valois FX, Sansonetti PJ. Tracking bacterial pathogens with genetically-encoded reporters. FEBS Lett. 2014;588: 2428–2436. doi: 10.1016/j.febslet.2014.05.022 - DOI - PubMed
-
- Charpentier X, Oswald E. Identification of the secretion and translocation domain of the enteropathogenic and enterohemorrhagic Escherichia coli effector Cif, using TEM-1 beta-lactamase as a new fluorescence-based reporter. J Bacteriol. 2004;186: 5486–5495. doi: 10.1128/JB.186.16.5486-5495.2004 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
