

14-3-3 Proteins in Brain Development: Neurogenesis, Neuronal Migration and Neuromorphogenesis
- PMID: 29075177
- PMCID: PMC5643407
- DOI: 10.3389/fnmol.2017.00318
14-3-3 Proteins in Brain Development: Neurogenesis, Neuronal Migration and Neuromorphogenesis
Abstract
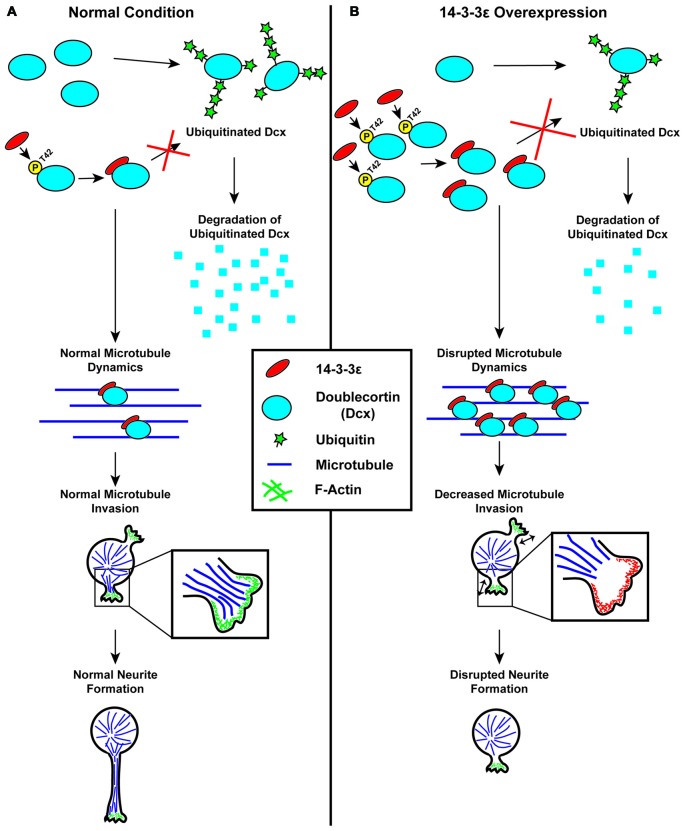
The 14-3-3 proteins are a family of highly conserved, multifunctional proteins that are highly expressed in the brain during development. Cumulatively, the seven 14-3-3 isoforms make up approximately 1% of total soluble brain protein. Over the last decade, evidence has accumulated implicating the importance of the 14-3-3 protein family in the development of the nervous system, in particular cortical development, and have more recently been recognized as key regulators in a number of neurodevelopmental processes. In this review we will discuss the known roles of each 14-3-3 isoform in the development of the cortex, their relation to human neurodevelopmental disorders, as well as the challenges and questions that are left to be answered. In particular, we focus on the 14-3-3 isoforms and their involvement in the three key stages of cortical development; neurogenesis and differentiation, neuronal migration and neuromorphogenesis and synaptogenesis.
Keywords: 14-3-3 proteins; neurite growth; neurite initiation; neurodevelopmental disorders; neurogenesis; neuromorphogenesis; neuronal migration; synaptogenesis.
Figures
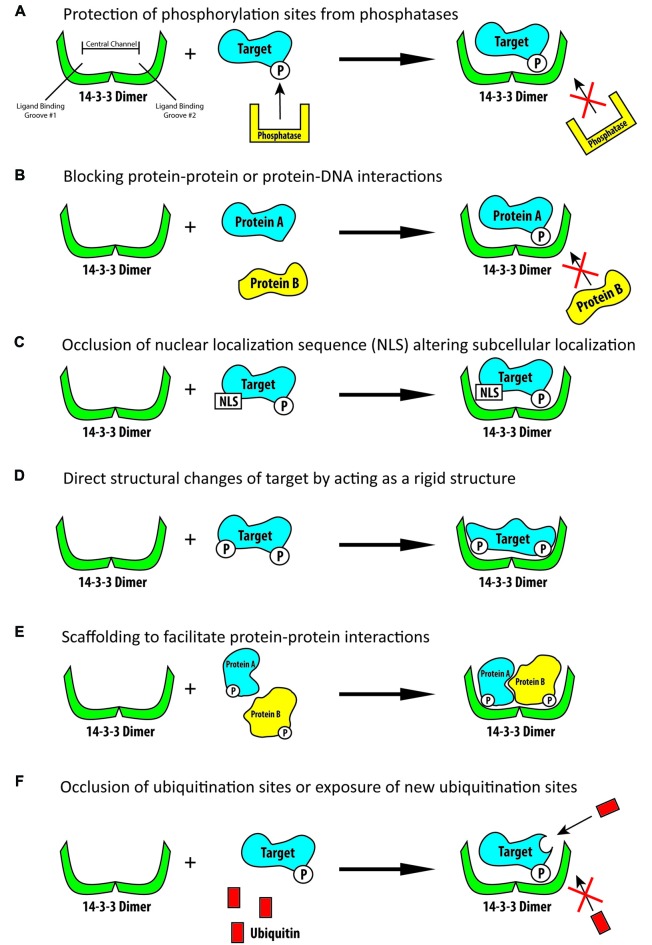
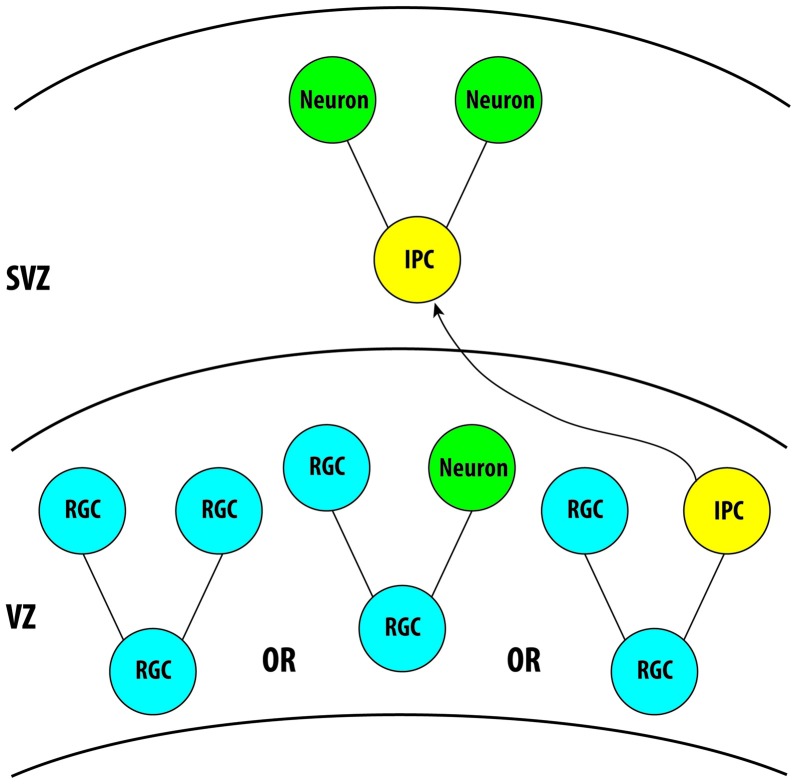
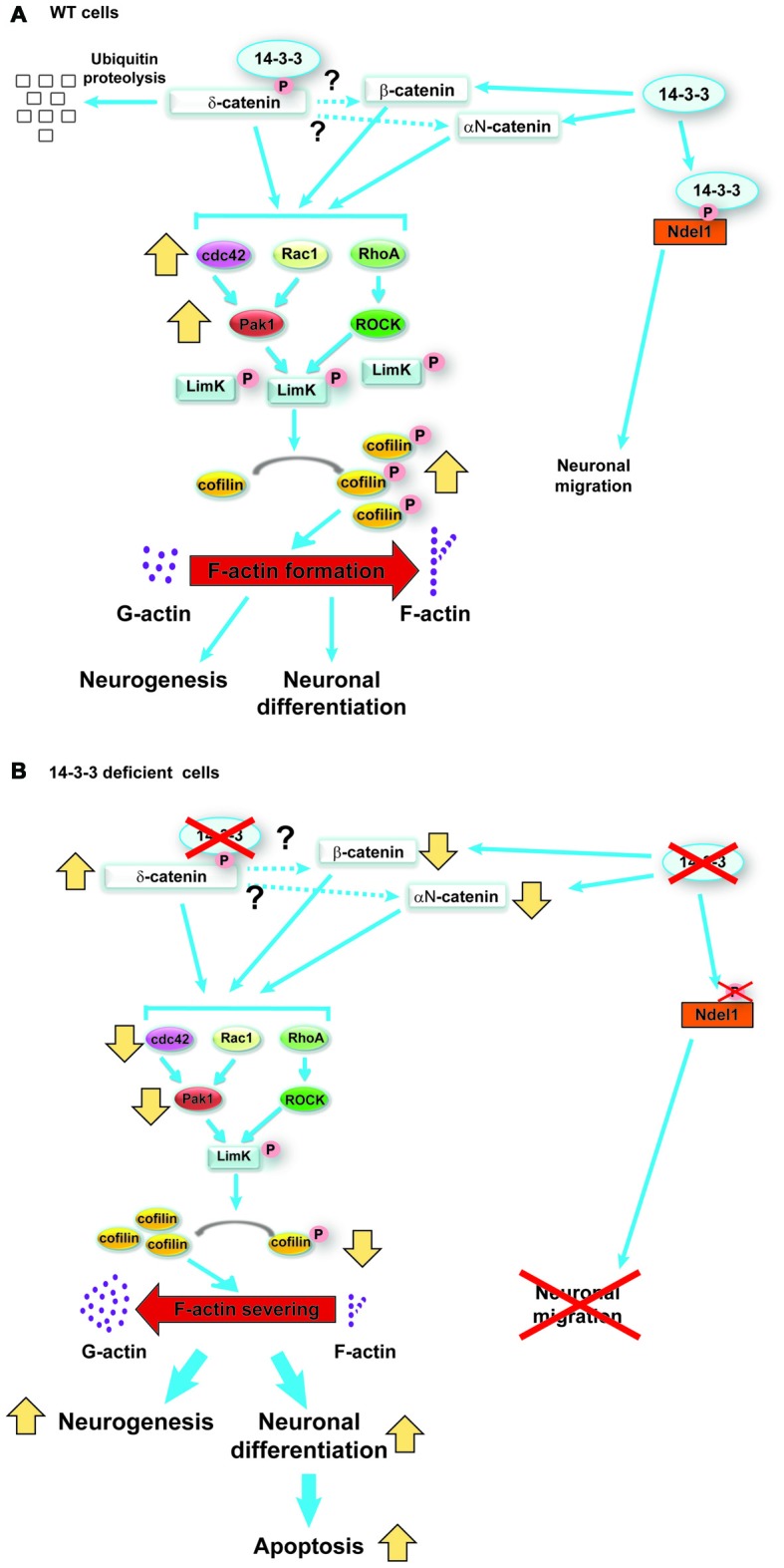
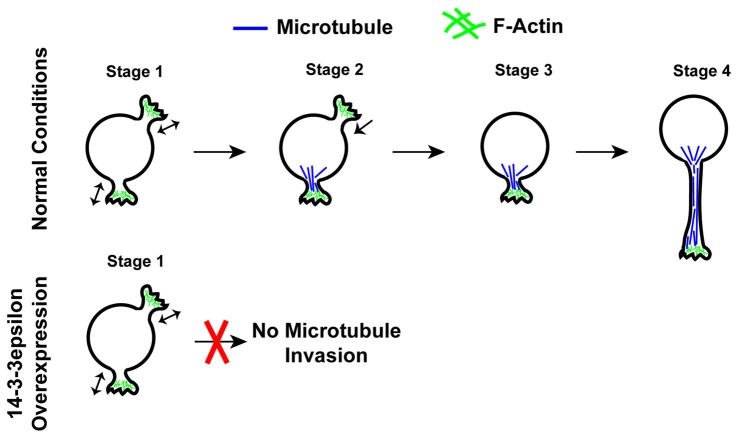
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources