

The flipflop orphan genes are required for limb bud eversion in the Tribolium embryo
- PMID: 29075305
- PMCID: PMC5649079
- DOI: 10.1186/s12983-017-0234-9
The flipflop orphan genes are required for limb bud eversion in the Tribolium embryo
Abstract
Background: Unlike Drosophila but similar to other arthropod and vertebrate embryos, the flour beetle Tribolium castaneum develops everted limb buds during embryogenesis. However, the molecular processes directing the evagination of epithelia are only poorly understood.
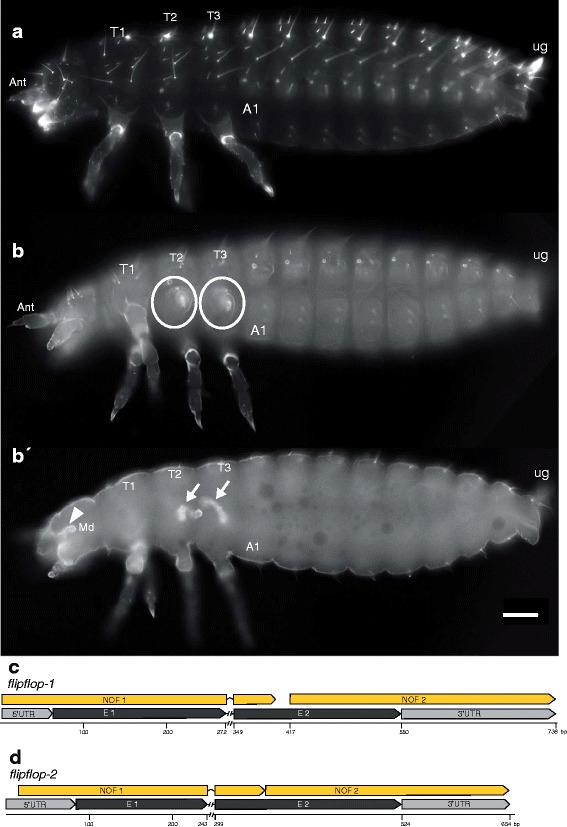
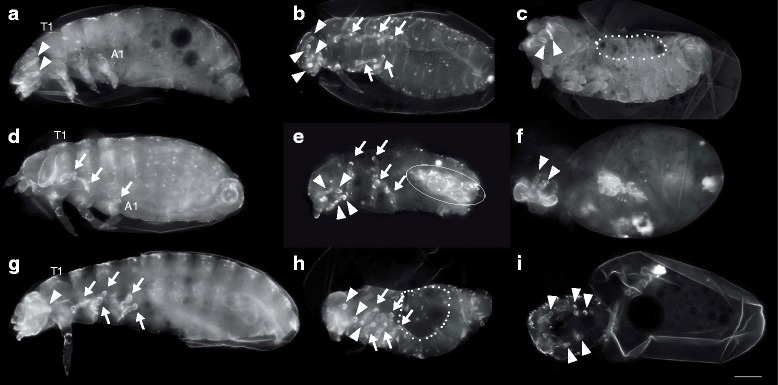
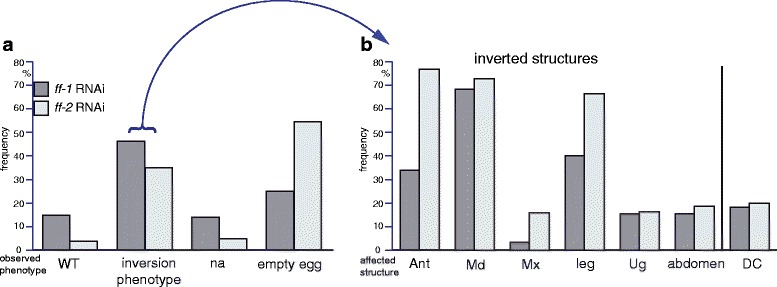
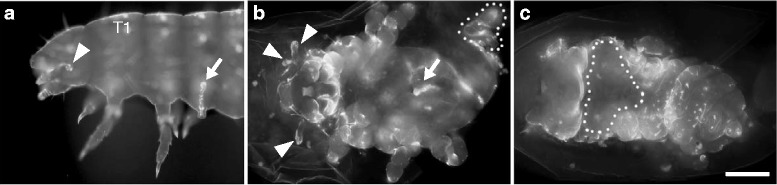
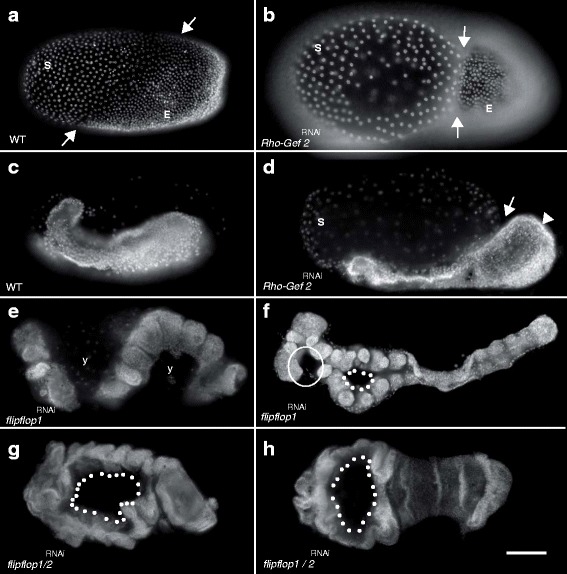
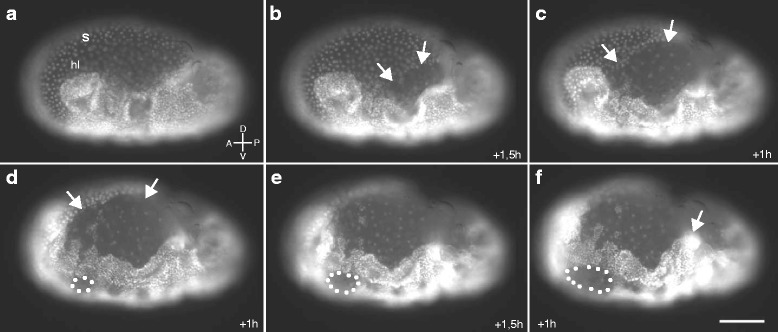
Results: Here we show that the newly discovered genes Tc-flipflop1 and Tc-flipflop2 are involved in regulating the directional budding of appendages. RNAi-knockdown of Tc-flipflop results in a variety of phenotypic traits. Most prominently, embryonic limb buds frequently grow inwards rather than out, leading to the development of inverted appendages inside the larval body. Moreover, affected embryos display dorsal closure defects. The Tc-flipflop genes are evolutionarily non-conserved, and their molecular function is not evident. We further found that Tc-RhoGEF2, a highly-conserved gene known to be involved in actomyosin-dependent cell movement and cell shape changes, shows a Tc-flipflop-like RNAi-phenotype.
Conclusions: The similarity of the inverted appendage phenotype in both the flipflop- and the RhoGEF2 RNAi gene knockdown led us to conclude that the Tc-flipflop orphan genes act in a Rho-dependent pathway that is essential for the early morphogenesis of polarised epithelial movements. Our work describes one of the few examples of an orphan gene playing a crucial role in an important developmental process.
Keywords: Appendage formation; Epithelial morphogenesis; Evagination; Orphan flipflop gene; PCP; RhoGEF2; Tissue folding; Tribolium castaneum.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures


References
-
- Barrett K, Leptin M, Settleman J. The Rho GTPase and a putative RhoGEF mediate a signaling pathway for the cell shape changes in Drosophila gastrulation. Cell. 1997;91:905–15. - PubMed
-
- Beermann A, Jay DG, Beeman RW, Hülskamp M, Tautz D, Jürgens G. The Short antennae gene of Tribolium is required for limb development and encodes the orthologue of the Drosophila Distal-less protein. Development. 2001;128:287–97. - PubMed
-
- Brown, SJ, Shippy, TD, Miller, S, Bolognesi, R, Beeman, RW, Lorenzen, MD, Bucher, G, Wimmer, EA & Klingler, M. The Red Flour Beetle, Tribolium castaneum (Coleoptera): A Model for Studies of Development and Pest Biology. Cold Spring Harb Protoc 2009, doi:https://doi.org/10.1101/pdb.emo126. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
