

Recent evidence that TADs and chromatin loops are dynamic structures
- PMID: 29077530
- PMCID: PMC5990973
- DOI: 10.1080/19491034.2017.1389365
Recent evidence that TADs and chromatin loops are dynamic structures
Abstract
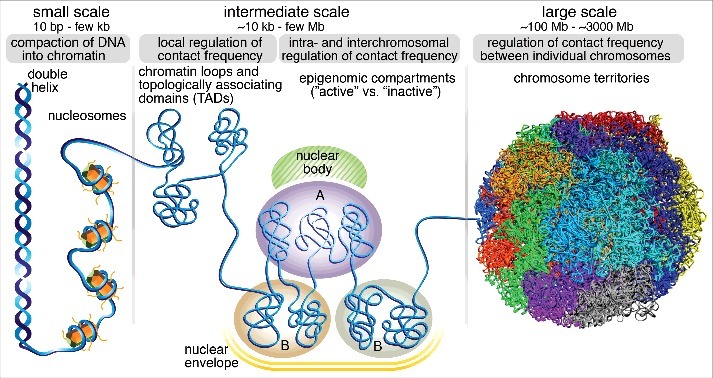
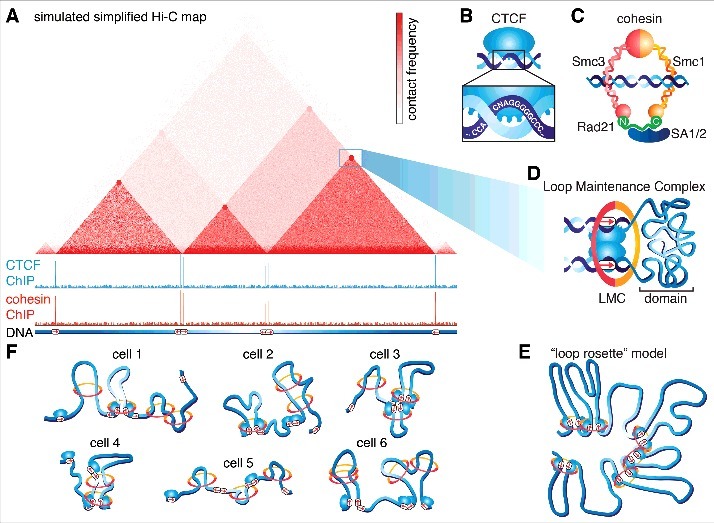
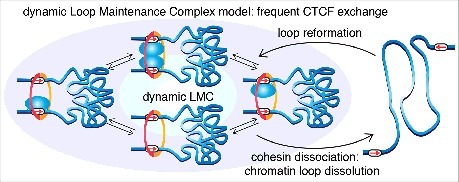
Mammalian genomes are folded into spatial domains, which regulate gene expression by modulating enhancer-promoter contacts. Here, we review recent studies on the structure and function of Topologically Associating Domains (TADs) and chromatin loops. We discuss how loop extrusion models can explain TAD formation and evidence that TADs are formed by the ring-shaped protein complex, cohesin, and that TAD boundaries are established by the DNA-binding protein, CTCF. We discuss our recent genomic, biochemical and single-molecule imaging studies on CTCF and cohesin, which suggest that TADs and chromatin loops are dynamic structures. We highlight complementary polymer simulation studies and Hi-C studies employing acute depletion of CTCF and cohesin, which also support such a dynamic model. We discuss the limitations of each approach and conclude that in aggregate the available evidence argues against stable loops and supports a model where TADs are dynamic structures that continually form and break throughout the cell cycle.
Keywords: 3D genome; CTCF; FRAP; chromatin loops; cohesin; dynamics; loop extrusion; modeling; single-molecule imaging; topological domains.
Figures
Comment on
- Extra view to: AS Hansen, I Pustova, C Cattoglio, R Tjian, X. Darzacq. CTCF and cohesin regulate chromatin loop stability with distinct dynamics. Elife. 2017; 6; . doi:https://doi.org/10.7554/eLife.2577610.7554/eLife.25776 doi: 10.7554/eLife.25776
References
-
- Merkenschlager M, Nora EP. CTCF and cohesin in genome folding and transcriptional gene regulation. Annu Rev Genomics Hum Genet. [Internet] 2017;17:1–27. Available from: http://www.ncbi.nlm.nih.gov/pubmed/27089971 - PubMed
-
- Ruiz-Velasco M, Zaugg JB. Structure meets function: how chromatin organisation conveys functionality. Curr Opin Syst Biol. [Internet] 2017;1:129–36. Available from: http://www.sciencedirect.com/science/article/pii/S2452310017300173 https://doi.org/ 10.1016/j.coisb.2017.01.003. - DOI
-
- Nora EP, Lajoie BR, Schulz EG, Giorgetti L, Okamoto I, Servant N, Piolot T, van Berkum NL, Meisig J, Sedat J, et al.. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature. 2012;485:381–5. PMID:22495304 https://doi.org/ 10.1038/nature11049. - DOI - PMC - PubMed
-
- Dixon JR, Selvaraj S, Yue F, Kim A, Li Y, Shen Y, Hu M, Liu JS, Ren B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. [Internet] 2012;485:376–80. Available from: https://doi.org/10.1038/nature11082%5Cnhttp://www.nature.com/doifinder/1... https://doi.org/ 10.1038/nature11082. - DOI - PMC - PubMed
-
- Lieberman-aiden E, Berkum NL Van, Williams L, Imakaev M, Ragoczy T, Telling A, Amit I, Lajoie BR, Sabo PJ, Dorschner MO, et al.. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. (80-) 2009;326:289–93.https://doi.org/ 10.1126/science.1181369. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous