

Lack of BACE1 S-palmitoylation reduces amyloid burden and mitigates memory deficits in transgenic mouse models of Alzheimer's disease
- PMID: 29078331
- PMCID: PMC5692556
- DOI: 10.1073/pnas.1708568114
Lack of BACE1 S-palmitoylation reduces amyloid burden and mitigates memory deficits in transgenic mouse models of Alzheimer's disease
Abstract
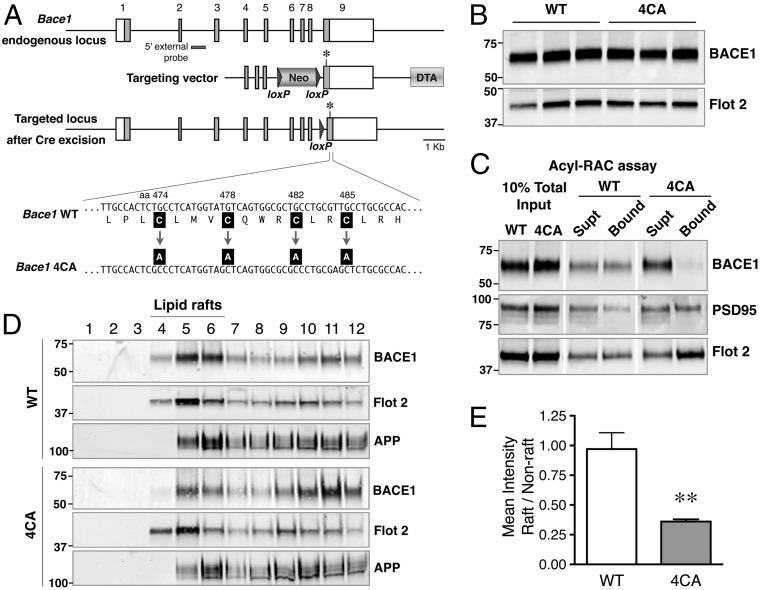
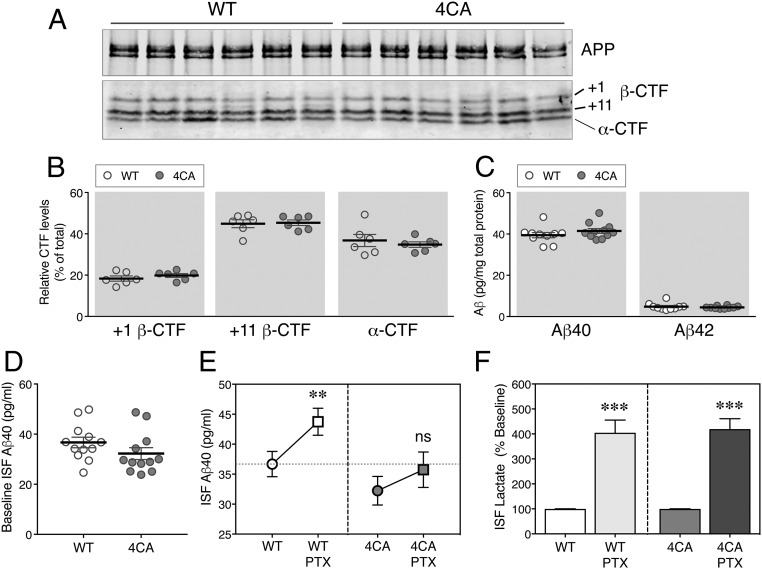
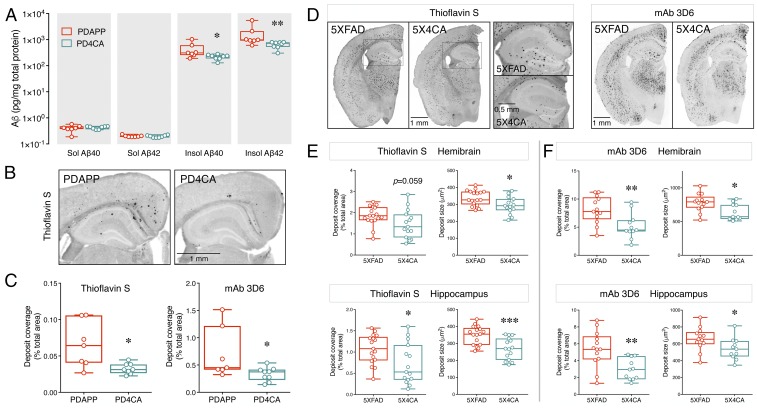
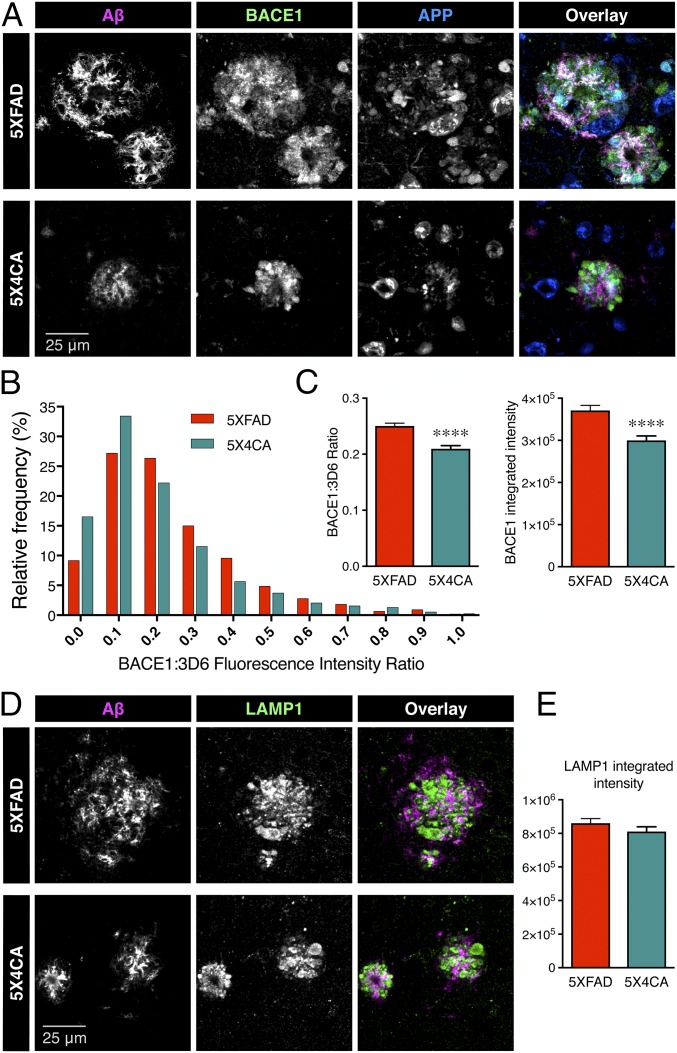
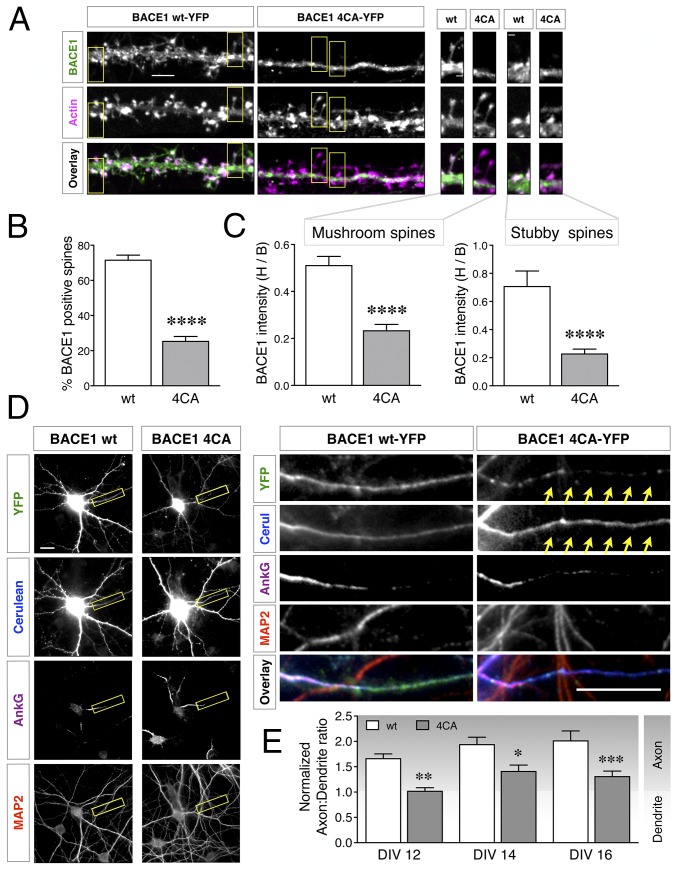
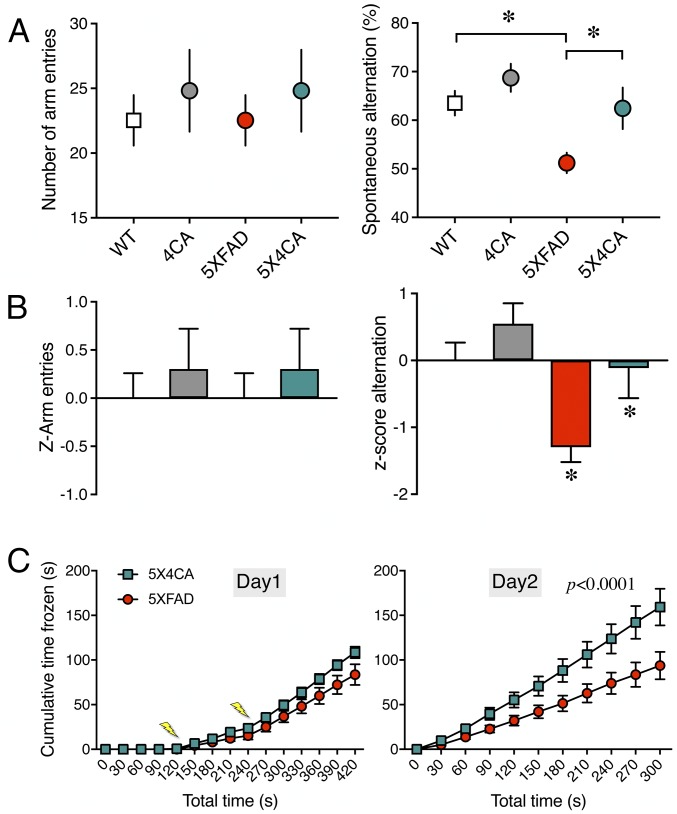
Alzheimer's disease (AD) is a devastating neurodegenerative disorder characterized by pathological brain lesions and a decline in cognitive function. β-Amyloid peptides (Aβ), derived from proteolytic processing of amyloid precursor protein (APP), play a central role in AD pathogenesis. β-Site APP cleaving enzyme 1 (BACE1), the transmembrane aspartyl protease which initiates Aβ production, is axonally transported in neurons and accumulates in dystrophic neurites near cerebral amyloid deposits in AD. BACE1 is modified by S-palmitoylation at four juxtamembrane cysteine residues. S-palmitoylation is a dynamic posttranslational modification that is important for trafficking and function of several synaptic proteins. Here, we investigated the in vivo significance of BACE1 S-palmitoylation through the analysis of knock-in mice with cysteine-to-alanine substitution at the palmitoylated residues (4CA mice). BACE1 expression, as well as processing of APP and other neuronal substrates, was unaltered in 4CA mice despite the lack of BACE1 S-palmitoylation and reduced lipid raft association. Whereas steady-state Aβ levels were similar, synaptic activity-induced endogenous Aβ production was not observed in 4CA mice. Furthermore, we report a significant reduction of cerebral amyloid burden and BACE1 accumulation in dystrophic neurites in the absence of BACE1 S-palmitoylation in mouse models of AD amyloidosis. Studies in cultured neurons suggest that S-palmitoylation is required for dendritic spine localization and axonal targeting of BACE1. Finally, the lack of BACE1 S-palmitoylation mitigates cognitive deficits in 5XFAD mice. Using transgenic mouse models, these results demonstrate that intrinsic posttranslational S-palmitoylation of BACE1 has a significant impact on amyloid pathogenesis and the consequent cognitive decline.
Keywords: 5XFAD; PDAPP; axonal transport; dystrophic neurite; neurodegeneration.
Published under the PNAS license.
Conflict of interest statement
Conflict of interest statement: D.M.H. is cofounder of C2N Diagnostics, LLC, serves on the scientific advisory board of C2N Diagnostics, and consults for Eli Lilly, AbbVie, GlaxoSmithKline, Genentech, Proclara Biosciences, and Denali.
Figures
References
-
- Mullan M, et al. A pathogenic mutation for probable Alzheimer’s disease in the APP gene at the N-terminus of beta-amyloid. Nat Genet. 1992;1:345–347. - PubMed
-
- Citron M, et al. Mutation of the beta-amyloid precursor protein in familial Alzheimer’s disease increases beta-protein production. Nature. 1992;360:672–674. - PubMed
-
- Cai XD, Golde TE, Younkin SG. Release of excess amyloid beta protein from a mutant amyloid beta protein precursor. Science. 1993;259:514–516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
