

Bacteriorhodopsin-like channelrhodopsins: Alternative mechanism for control of cation conductance
- PMID: 29078348
- PMCID: PMC5692563
- DOI: 10.1073/pnas.1710702114
Bacteriorhodopsin-like channelrhodopsins: Alternative mechanism for control of cation conductance
Abstract
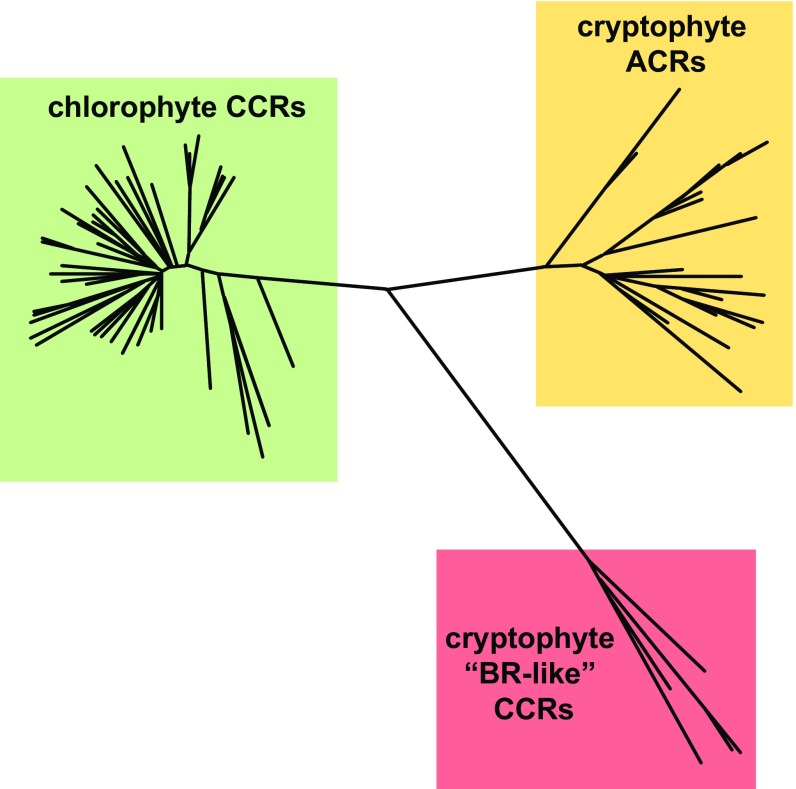
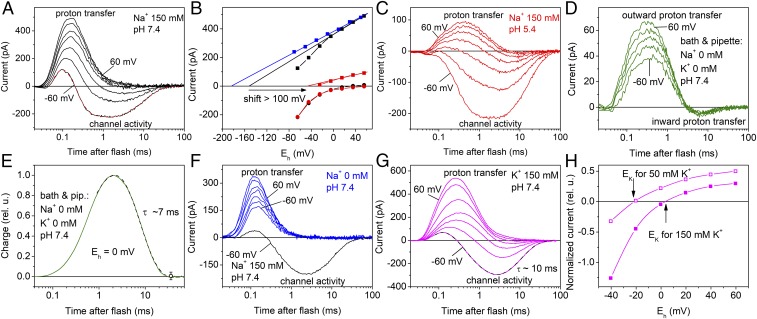
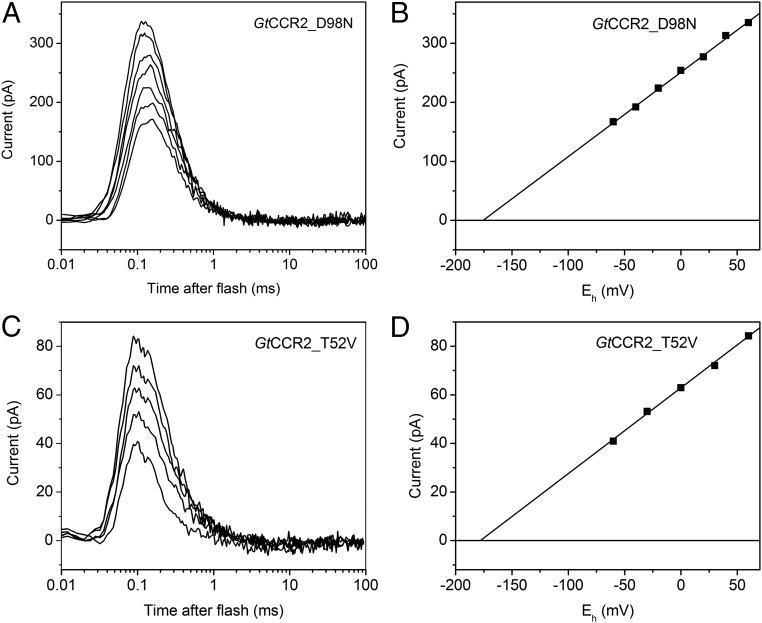
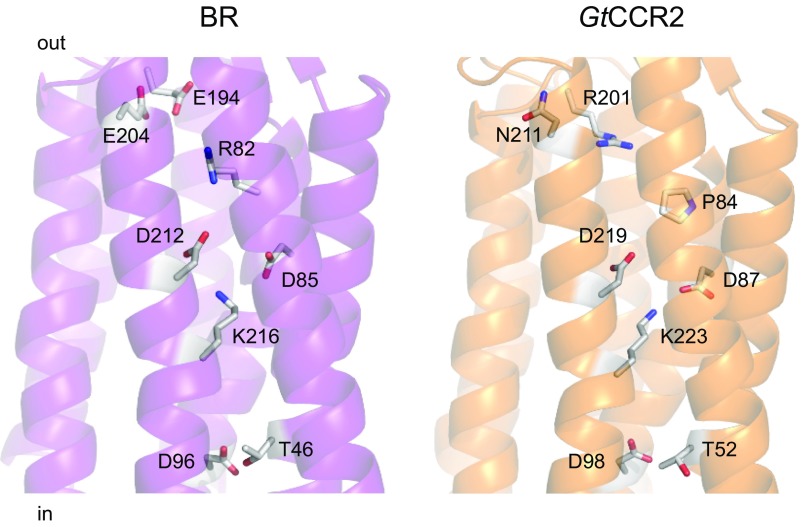
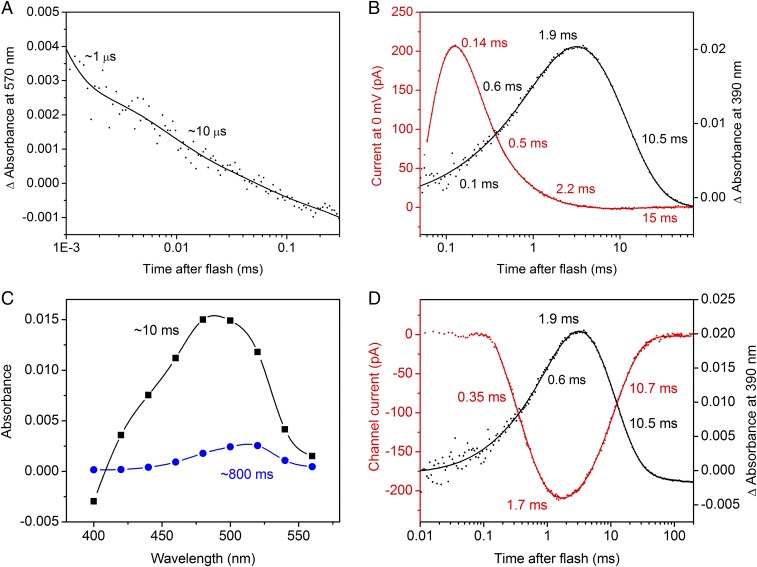
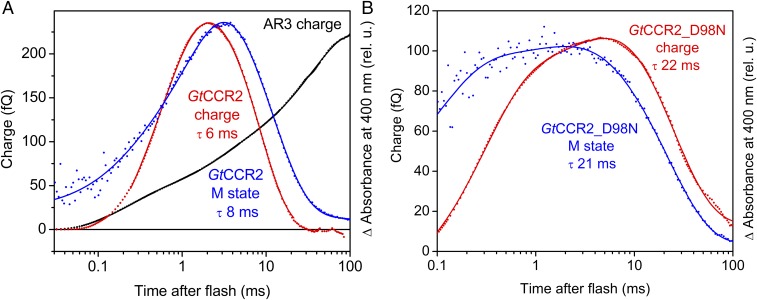
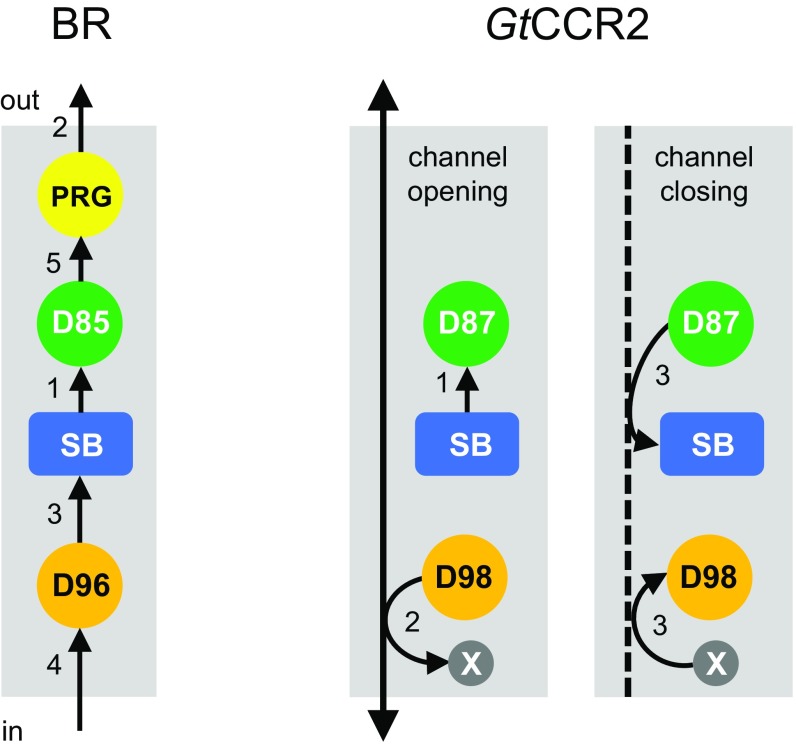
The recently discovered cation-conducting channelrhodopsins in cryptophyte algae are far more homologous to haloarchaeal rhodopsins, in particular the proton pump bacteriorhodopsin (BR), than to earlier known channelrhodopsins. They uniquely retain the two carboxylate residues that define the vectorial proton path in BR in which Asp-85 and Asp-96 serve as acceptor and donor, respectively, of the photoactive site Schiff base (SB) proton. Here we analyze laser flash-induced photocurrents and photochemical conversions in Guillardia theta cation channelrhodopsin 2 (GtCCR2) and its mutants. Our results reveal a model in which the GtCCR2 retinylidene SB chromophore rapidly deprotonates to the Asp-85 homolog, as in BR. Opening of the cytoplasmic channel to cations in GtCCR2 requires the Asp-96 homolog to be unprotonated, as has been proposed for the BR cytoplasmic channel for protons. However, reprotonation of the GtCCR2 SB occurs not from the Asp-96 homolog, but by proton return from the earlier protonated acceptor, preventing vectorial proton translocation across the membrane. In GtCCR2, deprotonation of the Asp-96 homolog is required for cation channel opening and occurs >10-fold faster than reprotonation of the SB, which temporally correlates with channel closing. Hence in GtCCR2, cation channel gating is tightly coupled to intramolecular proton transfers involving the same residues that define the vectorial proton path in BR.
Keywords: channelrhodopsins; ion transport; optogenetics; photocycle; proton transfers.
Published under the PNAS license.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nagel G, et al. Channelrhodopsin-1: A light-gated proton channel in green algae. Science. 2002;296:2395–2398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
