

Functional partnership between mGlu3 and mGlu5 metabotropic glutamate receptors in the central nervous system
- PMID: 29079293
- PMCID: PMC6263139
- DOI: 10.1016/j.neuropharm.2017.10.026
Functional partnership between mGlu3 and mGlu5 metabotropic glutamate receptors in the central nervous system
Abstract
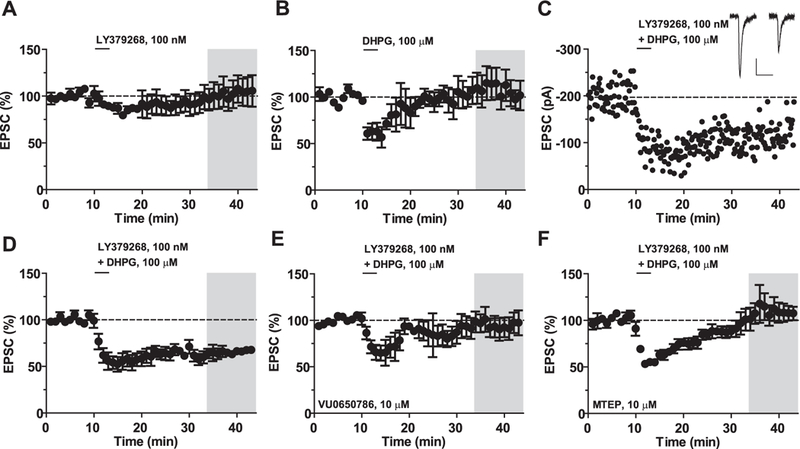
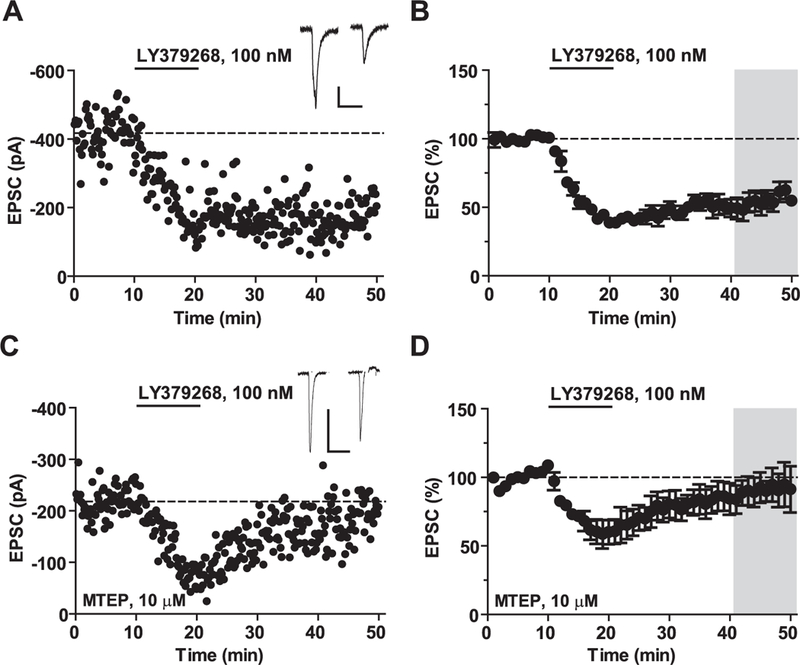
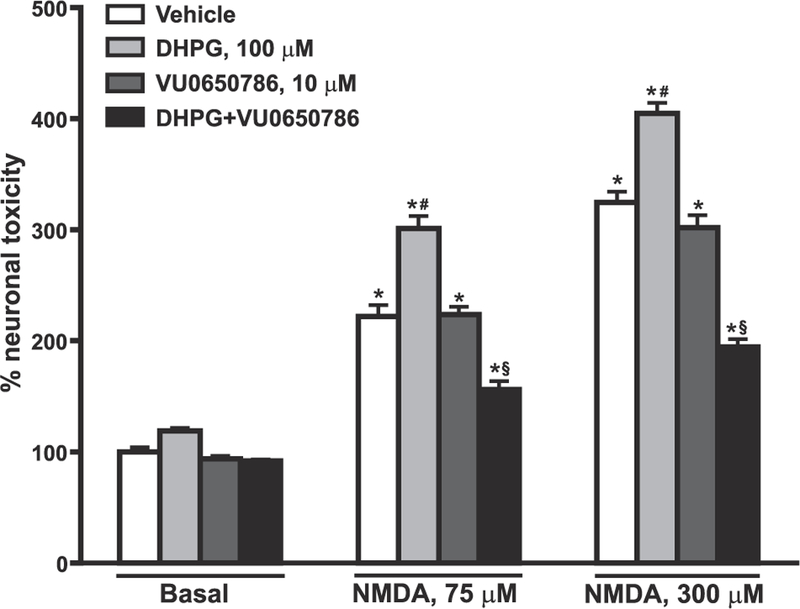
mGlu5 receptors are involved in mechanisms of activity-dependent synaptic plasticity, and are targeted by drugs developed for the treatment of CNS disorders. We report that mGlu3 receptors, which are traditionally linked to the control of neurotransmitter release, support mGlu5 receptor signaling in neurons and largely contribute to the robust mGlu5 receptor-mediated polyphosphoinositide hydrolysis in the early postnatal life. In cortical pyramidal neurons, mGlu3 receptor activation potentiated mGlu5 receptor-mediated somatic Ca2+ mobilization, and mGlu3 receptor-mediated long-term depression in the prefrontal cortex required the endogenous activation of mGlu5 receptors. The interaction between mGlu3 and mGlu5 receptors was also relevant to mechanisms of neuronal toxicity, with mGlu3 receptors shaping the influence of mGlu5 receptors on excitotoxic neuronal death. These findings shed new light into the complex role played by mGlu receptors in physiology and pathology, and suggest reconsideration of some of the current dogmas in the mGlu receptor field.
Keywords: G-protein βγ subunits; Long-term depression; Metabotropic glutamate receptors; Neurodevelopment; Neuronal death; Polyphosphoinositide hydrolysis; Receptor-receptor cross-talk; Synaptic plasticity.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
References
-
- Aghajanian GK, Marek GJ, 2000. Serotonin model of schizophrenia: emerging role of glutamate mechanisms. Brain Res. Brain Res. Rev 31, 302–312. - PubMed
-
- Alagarsamy S, Marino MJ, Rouse ST, Gereau RW 4th, Heinemann SF, Conn PJ, 1999. Activation of NMDA receptors reverses desensitization of mGluR5 in native and recombinant systems. Nat. Neurosci 2, 234–240. - PubMed
-
- Alagarsamy S, Rouse ST, Junge C, Hubert GW, Gutman D, Smith Y, Conn PJ, 2002. NMDA-induced phosphorylation and regulation of mGluR5. Pharmacol. Biochem. Behav 73, 299–306. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
