

Essentiality of Regulator of G Protein Signaling 6 and Oxidized Ca2+/Calmodulin-Dependent Protein Kinase II in Notch Signaling and Cardiovascular Development
- PMID: 29079565
- PMCID: PMC5721783
- DOI: 10.1161/JAHA.117.007038
Essentiality of Regulator of G Protein Signaling 6 and Oxidized Ca2+/Calmodulin-Dependent Protein Kinase II in Notch Signaling and Cardiovascular Development
Abstract
Background: Congenital heart defects are the most common birth defects worldwide. Although defective Notch signaling is the major cause of mouse embryonic death from cardiovascular defects, how Notch signaling is regulated during embryonic vasculogenesis and heart development is poorly understood.
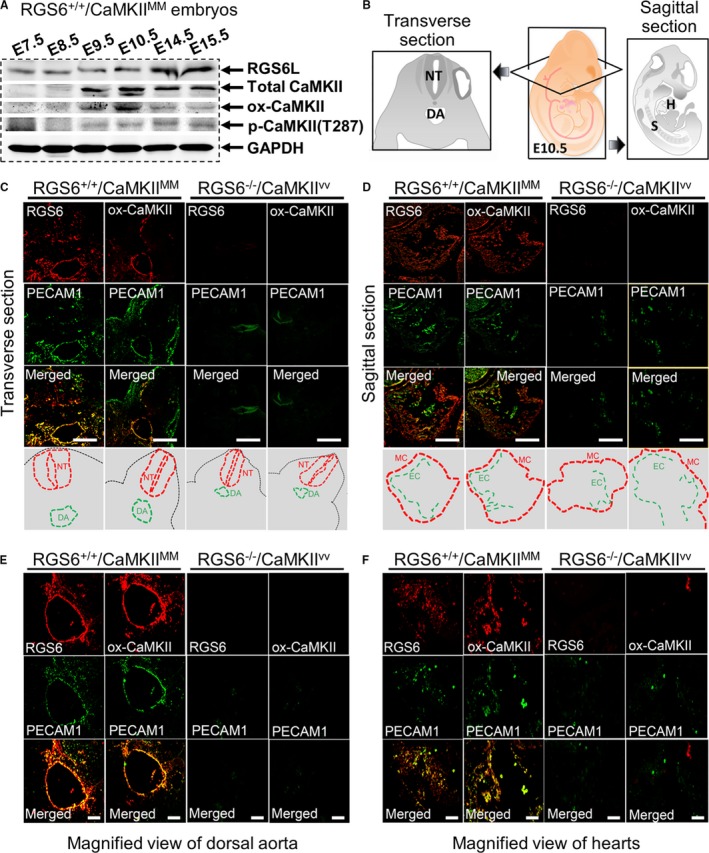
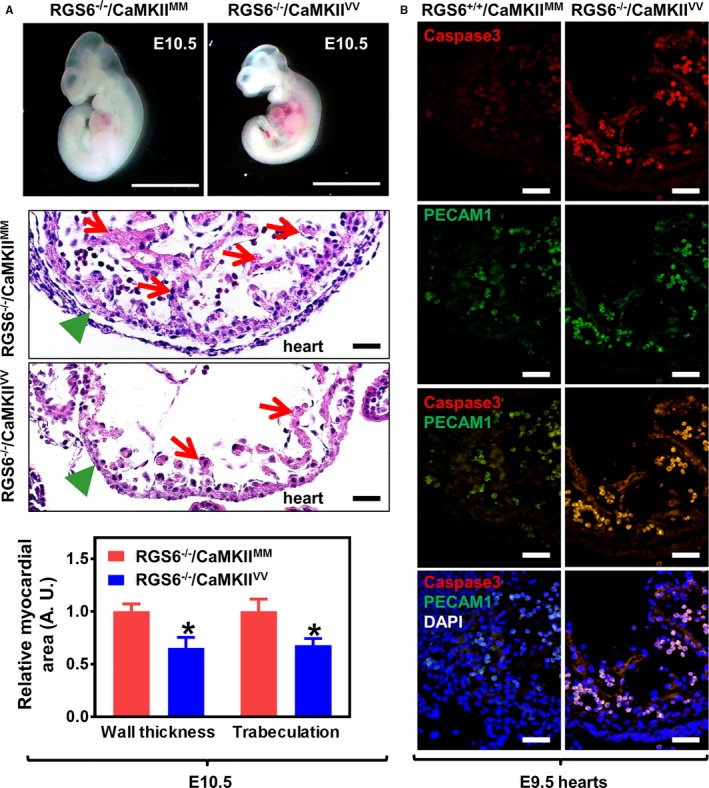
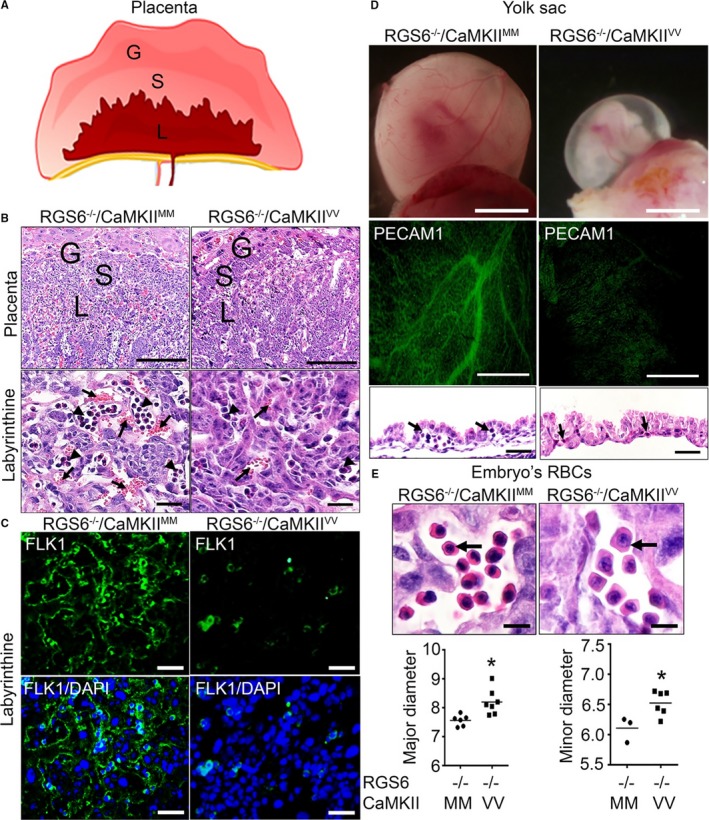
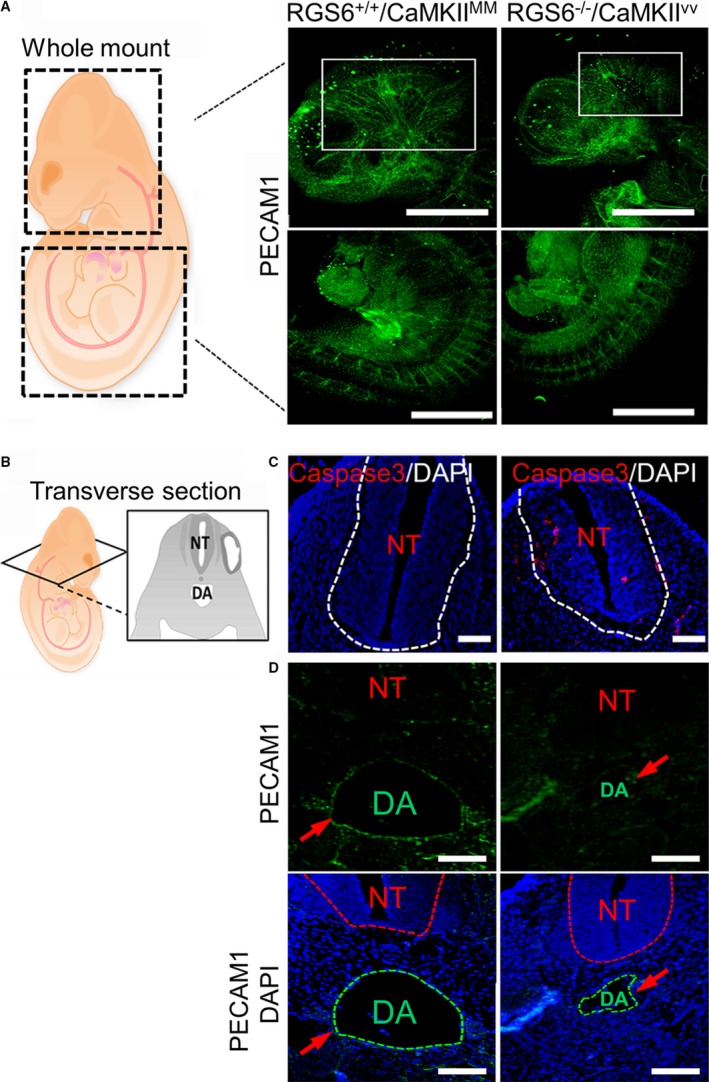
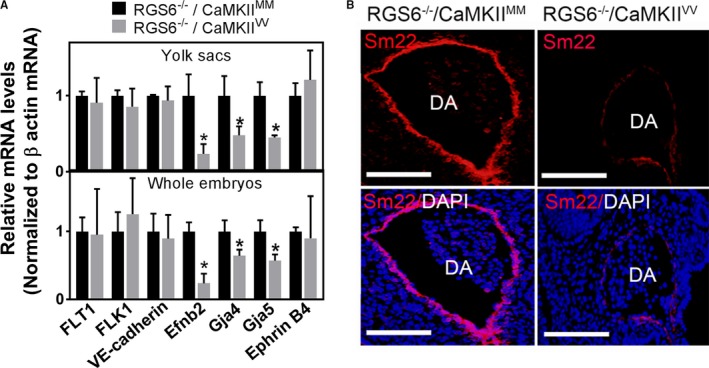
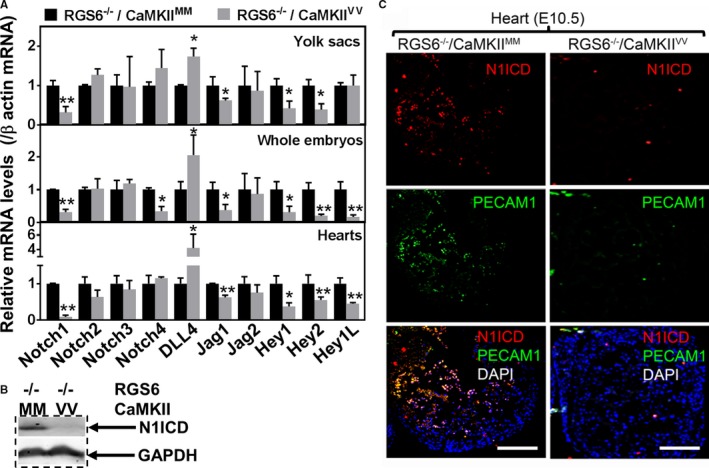
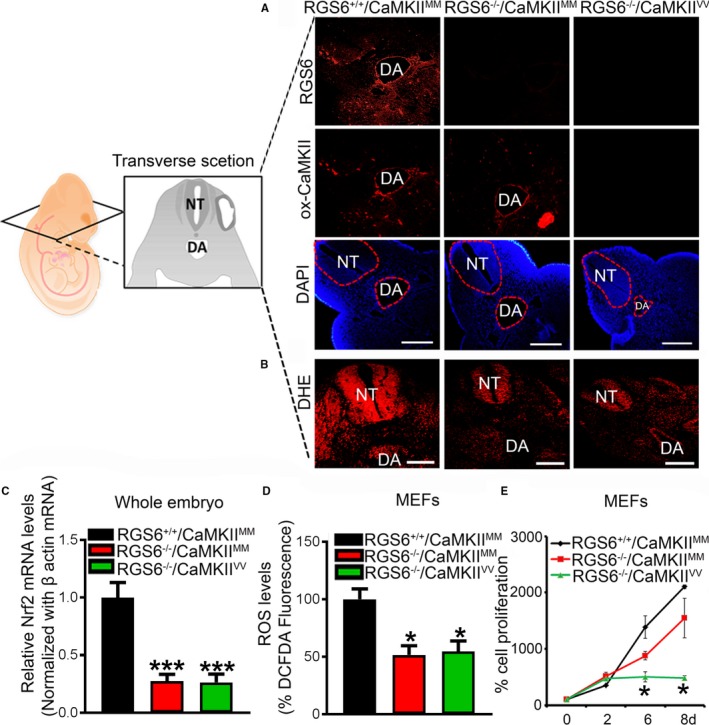
Methods and results: Regulator of G protein signaling 6 (RGS6)-/-/Ca2+/calmodulin-dependent protein kinase II (CaMKII)VV double mutant mice were developed by crossing RGS6-/- mice with mice expressing an oxidation-resistant CaMKIIδ (CaMKIIVV), and the resulting embryonic defects/lethality were investigated using E7.5 to E15.5 embryos. While loss of either RGS6 or oxidized CaMKIIδ does not alter embryogenesis, their combined loss causes defective Notch signaling, severe cardiovascular defects, and embryonic lethality (≈E10.5-11.5). Embryos lacking RGS6 and expressing oxidation-resistant CaMKIIδ exhibit reduced myocardial wall thickness, abnormal trabeculation, and arterial specification defects. Double mutants show vascular remodeling defects, including reduced neurovascularization, delayed neural tube maturation, and small dorsal aortae. These striking cardiovascular defects were accompanied by placental and yolk sac defects in angiogenesis, hematopoiesis, and vascular remodeling similar to what is seen with defective Notch1 signaling. Double mutant hearts, embryos, and yolk sacs exhibit profound downregulation of Notch1, Jagged 1, and Notch downstream target genes Hey1, Hey2, and Hey1L as well as impaired Notch1 signaling in embryos/hearts.
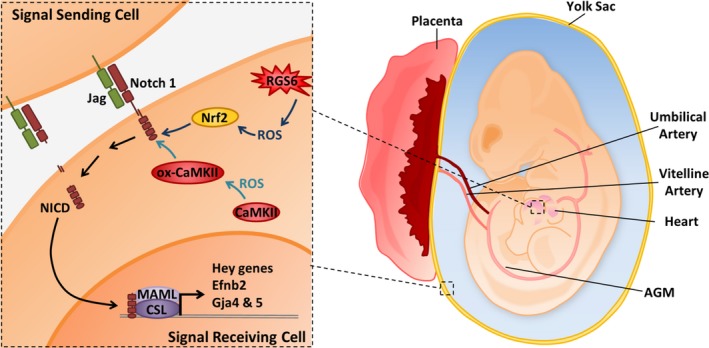
Conclusions: RGS6 and oxidized CaMKIIδ together function as novel critical upstream modulators of Notch signaling required for normal cardiovascular development and embryo survival. Their combined need indicates that they function in parallel pathways needed for Notch1 signaling in yolk sac, placenta and embryos. Thus, dysregulated embryonic RGS6 expression and oxidative activation of CaMKII may potentially contribute to congenital heart defects.
Keywords: Notch signaling; Regulator of G protein signaling 6; cardiovascular development; embryonic lethality; oxidized Ca2+/calmodulin‐dependent protein kinase II.
© 2017 The Authors. Published on behalf of the American Heart Association, Inc., by Wiley.
Figures
References
-
- Hoyert DL, Mathews TJ, Menacker F, Strobino DM, Guyer B. Annual summary of vital statistics: 2004. Pediatrics. 2006;117:168–183. - PubMed
-
- Bruneau BG. The developmental genetics of congenital heart disease. Nature. 2008;451:943–948. - PubMed
-
- Niessen K, Karsan A. Notch signaling in cardiac development. Circ Res. 2008;102:1169–1181. - PubMed
-
- Brand T. Heart development: molecular insights into cardiac specification and early morphogenesis. Dev Biol. 2003;258:1–19. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
