

Hypoxia modulates the purine salvage pathway and decreases red blood cell and supernatant levels of hypoxanthine during refrigerated storage
- PMID: 29079593
- PMCID: PMC5792281
- DOI: 10.3324/haematol.2017.178608
Hypoxia modulates the purine salvage pathway and decreases red blood cell and supernatant levels of hypoxanthine during refrigerated storage
Abstract
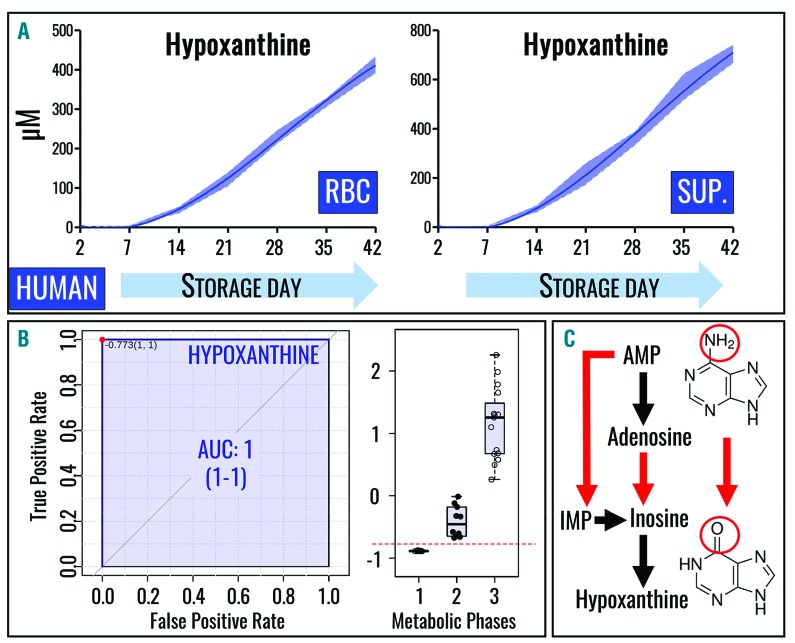
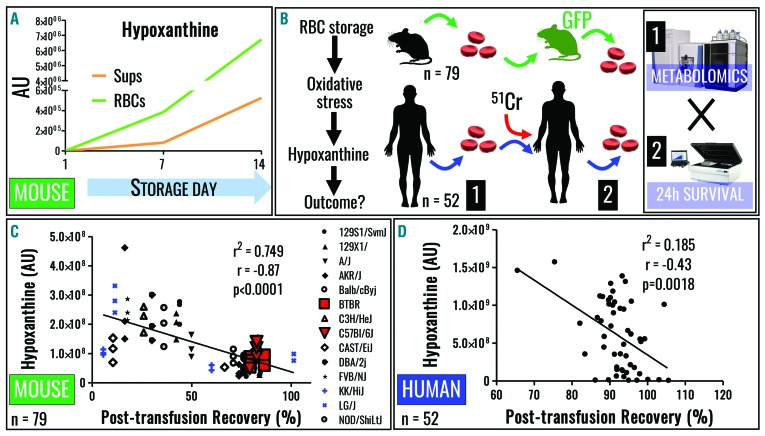
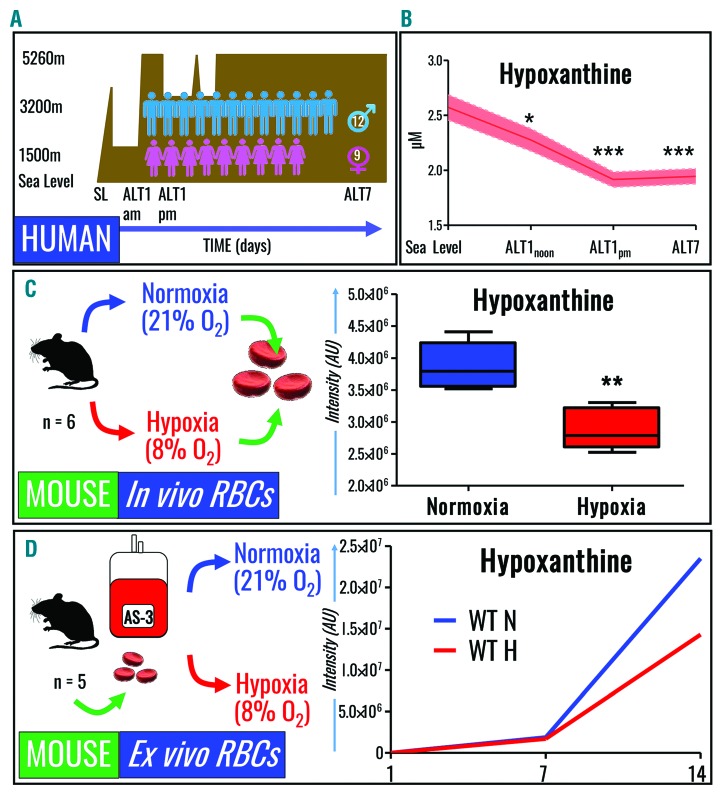
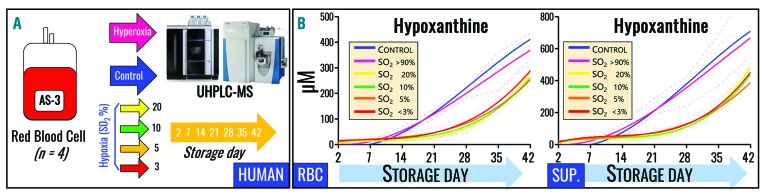
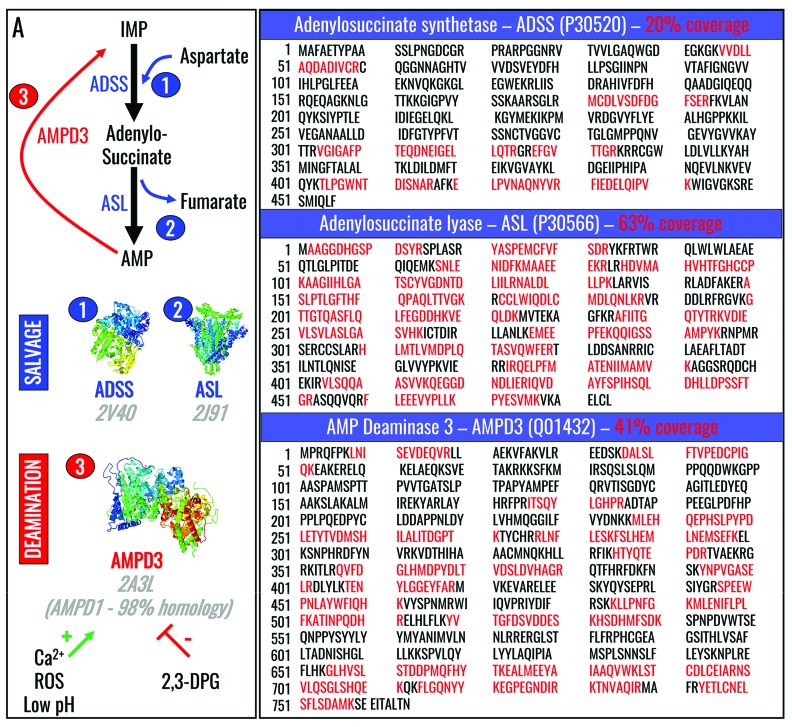
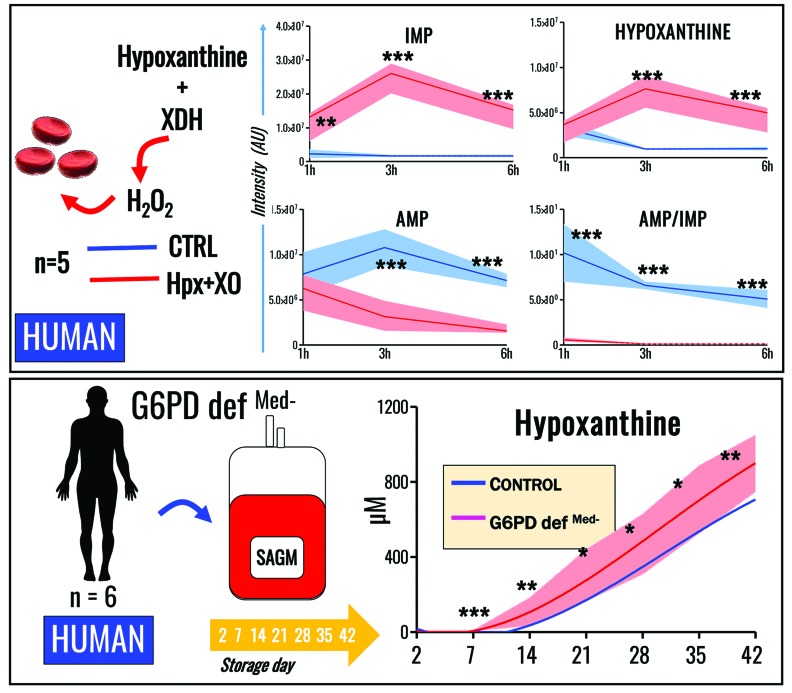
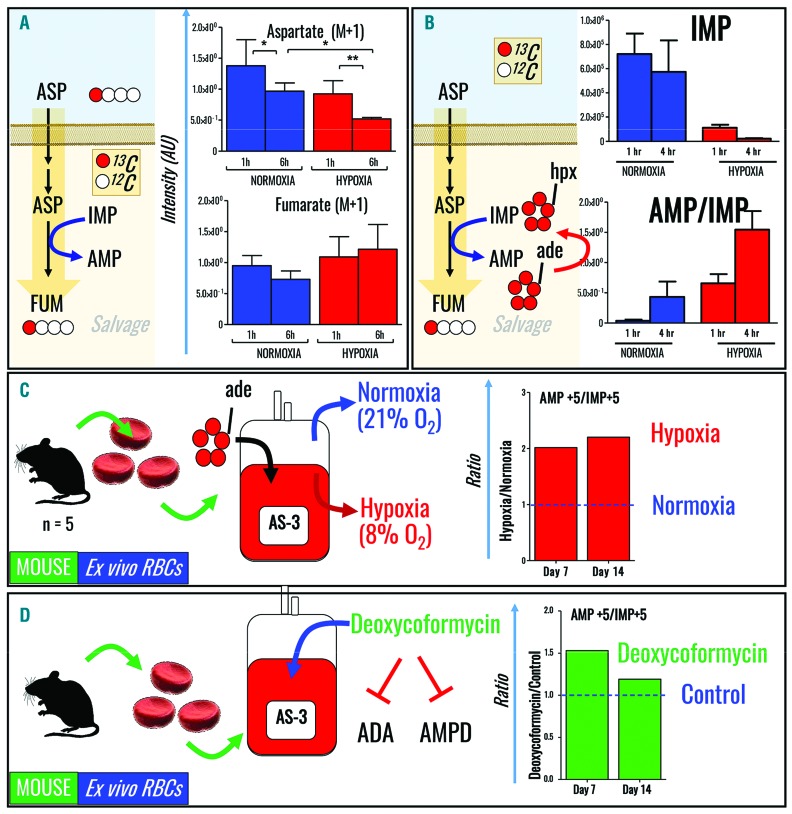
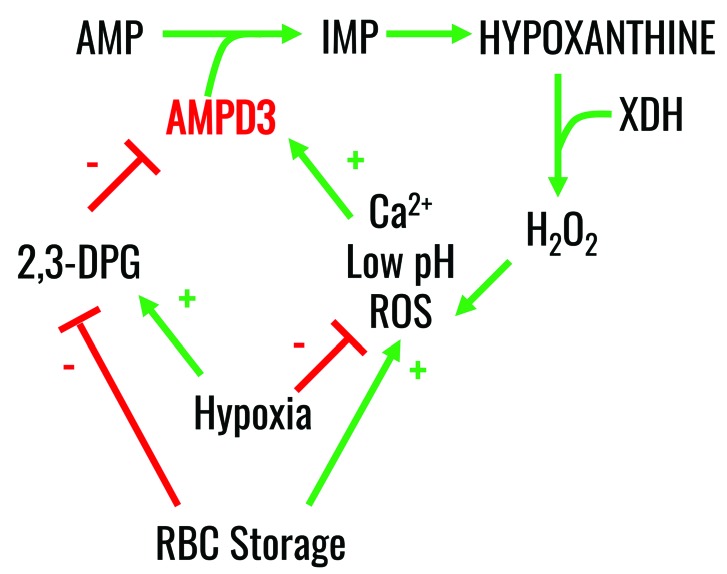
Hypoxanthine catabolism in vivo is potentially dangerous as it fuels production of urate and, most importantly, hydrogen peroxide. However, it is unclear whether accumulation of intracellular and supernatant hypoxanthine in stored red blood cell units is clinically relevant for transfused recipients. Leukoreduced red blood cells from glucose-6-phosphate dehydrogenase-normal or -deficient human volunteers were stored in AS-3 under normoxic, hyperoxic, or hypoxic conditions (with oxygen saturation ranging from <3% to >95%). Red blood cells from healthy human volunteers were also collected at sea level or after 1-7 days at high altitude (>5000 m). Finally, C57BL/6J mouse red blood cells were incubated in vitro with 13C1-aspartate or 13C5-adenosine under normoxic or hypoxic conditions, with or without deoxycoformycin, a purine deaminase inhibitor. Metabolomics analyses were performed on human and mouse red blood cells stored for up to 42 or 14 days, respectively, and correlated with 24 h post-transfusion red blood cell recovery. Hypoxanthine increased in stored red blood cell units as a function of oxygen levels. Stored red blood cells from human glucose-6-phosphate dehydrogenase-deficient donors had higher levels of deaminated purines. Hypoxia in vitro and in vivo decreased purine oxidation and enhanced purine salvage reactions in human and mouse red blood cells, which was partly explained by decreased adenosine monophosphate deaminase activity. In addition, hypoxanthine levels negatively correlated with post-transfusion red blood cell recovery in mice and - preliminarily albeit significantly - in humans. In conclusion, hypoxanthine is an in vitro metabolic marker of the red blood cell storage lesion that negatively correlates with post-transfusion recovery in vivo Storage-dependent hypoxanthine accumulation is ameliorated by hypoxia-induced decreases in purine deamination reaction rates.
Copyright© 2018 Ferrata Storti Foundation.
Figures
References
-
- D’Alessandro A, Kriebardis AG, Rinalducci S, et al. An update on red blood cell storage lesions, as gleaned through biochemistry and omics technologies. Transfusion. 2015;55(1):205–219. - PubMed
-
- Zimring JC. Widening our gaze of red blood storage haze: a role for metabolomics. Transfusion. 2015;55(6):1139–1142. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
