

Review
doi: 10.1104/pp.17.01272.
Epub 2017 Oct 27.
The Molecular Machinery of Chloroplast Division
Affiliations
- PMID: 29079653
- PMCID: PMC5761817
- DOI: 10.1104/pp.17.01272
Item in Clipboard
Review
The Molecular Machinery of Chloroplast Division
Plant Physiol.
2018 Jan.
Abstract
Recent studies advance understanding of the mechanisms, spatial control, and regulation of chloroplast division, but many questions remain.
Figures
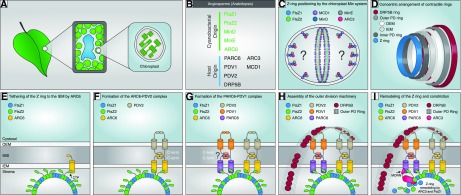
Working model of the positioning, assembly, and dynamics of the chloroplast division machinery in angiosperms based primarily on studies in Arabidopsis but informed by studies in C. merolae and other organisms. A, Diagram showing chloroplasts within a leaf mesophyll cell. B, Cyanobacterial (endosymbiotic) or host (eukaryotic) origin of chloroplast division components in angiosperms. C, FtsZ1 and FtsZ2 self-assemble as dynamic heteropolymers, possibly with mixed stoichiometry (Olson et al., 2010; Chen et al., 2017); protofilaments may possibly associate laterally to form the Z ring at the division site (Lutkenhaus and Du, 2017). Positioning of the Z ring is confined to the midzone by the chloroplast Min system, comprising ARC3, MCD1, MinD, and MinE, which inhibits Z-ring assembly at nondivision sites. ARC3 acts downstream of MinD and MinE as the direct inhibitor of Z-ring assembly (Zhang et al., 2013). MCD1, a transmembrane IEM protein, recruits MinD to the membrane (Nakanishi et al., 2009), where MinE is also colocalized (Miyagishima et al., 2011). ARC3 interacts with both MinD and MinE (Maple et al., 2007). Thus, we hypothesize that ARC3 forms a complex with MinD and MinE that is tethered to the membrane by MCD1. The exact localization pattern of ARC3 and the inhibitory mechanisms of ARC3 on Z-ring assembly are unclear (indicated by the question marks). The chloroplast Min-system components also localize partly to the division site (Shimada et al., 2004; Nakanishi et al., 2009; Miyagishima et al., 2011), where they may promote Z-ring remodeling during division (Johnson et al., 2015). D, Overview of the four contractile ring structures formed across the two envelope membranes. The composition of the inner PD ring is unknown and is not shown in further panels. The outer PD ring is synthesized by PDR1 (not shown) in the red alga C. merolae (Yoshida et al., 2010). The order of assembly based on studies in C. merolae is Z ring, inner PD ring, outer PD ring, DRP5B ring (Miyagishima et al., 2001, 2003). E to I, Stepwise assembly and dynamics of the division complex at the middle of the chloroplast. E, Tethering of the Z ring to the IEM is achieved mainly through interaction of the conserved FtsZ2 C-terminal peptide (CTP) with ARC6 (Maple et al., 2005), which probably stabilizes the Z ring and facilitates its assembly (Vitha et al., 2003; Johnson et al., 2013). F, ARC6 recruits PDV2 to the division site through direct interaction between their C-terminal IMS regions (Glynn et al., 2008). Dimerization of the cytosolic regions of two PDV2 molecules induces dimerization of two ARC6 molecules (Wang et al., 2017). G, PARC6 acts downstream of ARC6 to localize PDV1 to the division site through direct interaction between their C-terminal IMS regions (Glynn et al., 2009; Zhang et al., 2016). Based on ARC6-PDV2 studies (Wang et al., 2017), PDV1 dimerization might also promote PARC6 dimerization (indicated by question mark). H, In C. merolae, PDR1 (not shown) is recruited from the cytosol to construct the outer PD ring, composed of polyglucan fibrils (Yoshida et al., 2010). In Arabidopsis, PDV1 and PDV2 function together to recruit DRP5B from the cytosol (Miyagishima et al., 2006) to form the DRP5B ring (Gao et al., 2003; Miyagishima et al., 2003; Holtsmark et al., 2013). Coordination of the stromal Z ring and the cytosolic DRP5B ring is established through the ARC6-PDV2 and PARC6-PDV1 complexes. Whether the outer PD ring interacts with the PDV proteins is unclear. I, Remodeling of the Z ring and constriction. We speculate that PARC6 recruits ARC3 to the division site via interaction with the ARC3 MORN domain, enabling ARC3 to interact with FtsZ in the Z ring. The latter interaction may be facilitated by interaction of PARC6 with the FtsZ2 CTP (Zhang et al., 2016). As an FtsZ assembly inhibitor, ARC3 activation at the division site may promote Z-ring remodeling. MinD, MinE, and MCD1 also localize partly to the division site (Nakanishi et al., 2009; Miyagishima et al., 2011; not shown in I). Dynamic remodeling of the Z ring probably also depends on FtsZ1 (TerBush and Osteryoung, 2012; Yoshida et al., 2016; Terbush et al., 2018). OEM, Outer envelope membrane; IMS, intermembrane space; IEM, inner envelope membrane; MORN, membrane occupation and recognition nexus domain of ARC3; PD ring, plastid-dividing ring.
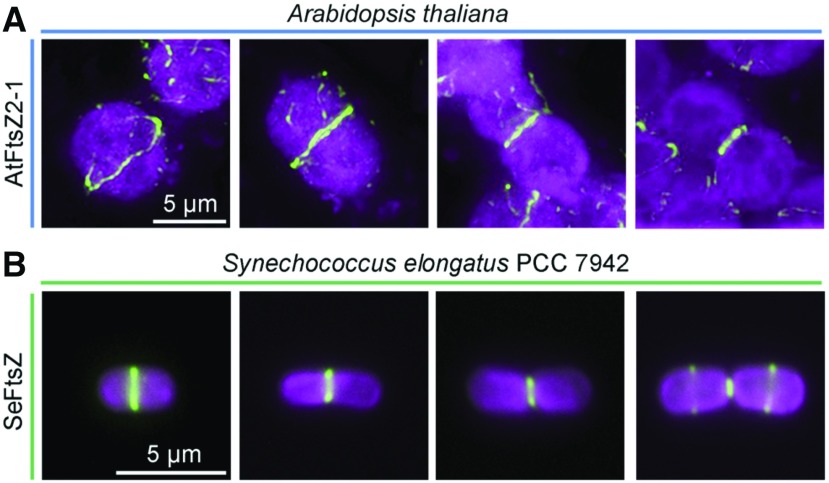
Immunofluorescence localization of the contractile Z ring in chloroplasts and cyanobacterial cells during division. A, Arabidopsis FtsZ2-1 (AtFtsZ2-1) detected with an anti-AtFtsZ2-1 antibody (McAndrew et al., 2001) in mesophyll cells of a fully expanded leaf obtained from a 3-week-old plant (Col-0). B, FtsZ in the cyanobacterium Synechococcus elongatus PCC 7942 (SeFtsZ) detected with an anti-Anabaena FtsZ antibody (Agrisera). Green, FtsZ; magenta, chlorophyll fluorescence. Bars, 5 μm.
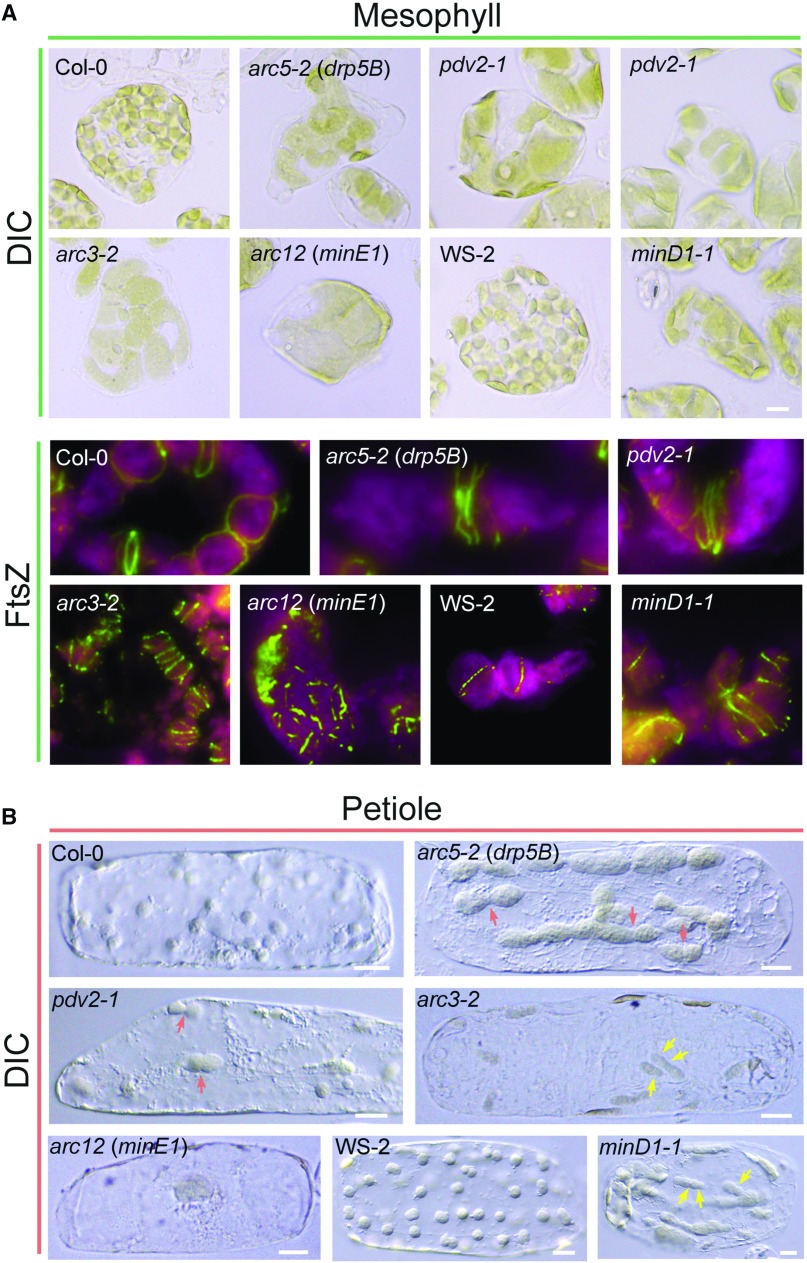
Chloroplast morphology and Z-ring localization patterns in Arabidopsis wild-type plants and various chloroplast division mutants. Although phenotypes vary, most mutants display reduced numbers of enlarged chloroplasts (Osteryoung and Pyke, 2014). A, Chloroplast morphology observed by differential interference contrast (DIC) microscopy (top) and Z-ring localization detected by immunofluorescence staining (bottom) in mesophyll cells. The minD1-1 mutant (Zhang et al., 2013) is in the WS-2 background; all others are in Col-0. Images of FtsZ localization were adapted from Miyagishima et al. (2006) (Col-0, pdv2-1 and arc5-2) and Zhang et al. (2013) (arc12 and minD1-1) with permission; copyright © 2006 and 2013 by the American Society of Plant Biologists. Green, FtsZ; magenta/red, chlorophyll fluorescence. B, Chloroplast morphology in cells isolated from leaf petioles. Petiole cells contain fewer chloroplasts than mesophyll cells, and the morphology phenotypes are more evident. Red arrows indicate dumbbell-shaped chloroplasts in pdv2-1 and arc5-2 mutants. Yellow arrows denote asymmetric division planes in arc3-2 and minD1-1 mutants. Bars, 10 μm.




References
-
- Abel WO, Knebel W, Koop H-U, Marienfeld JR, Quader H, Reski R, Schnepf E, Spörlein B (1989) A cytokinin-sensitive mutant of the moss, Physcomitrella patens, defective in chloroplast division. Protoplasma 152: 1–13
-
- Aldridge C, Møller SG (2005) The plastid division protein AtMinD1 is a Ca2+-ATPase stimulated by AtMinE1. J Biol Chem 280: 31673–31678 - PubMed
-
- Asano T, Yoshioka Y, Kurei S, Sakamoto W, Sodmergen, Machida Y (2004) A mutation of the CRUMPLED LEAF gene that encodes a protein localized in the outer envelope membrane of plastids affects the pattern of cell division, cell differentiation, and plastid division in Arabidopsis. Plant J 38: 448–459 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
