

γδ T Cells Are Required for the Induction of Sterile Immunity during Irradiated Sporozoite Vaccinations
- PMID: 29079696
- PMCID: PMC5698172
- DOI: 10.4049/jimmunol.1700314
γδ T Cells Are Required for the Induction of Sterile Immunity during Irradiated Sporozoite Vaccinations
Abstract
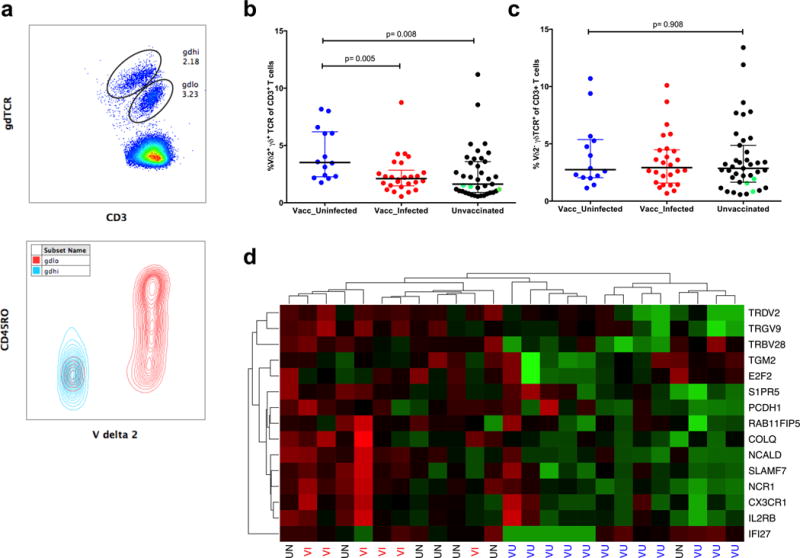
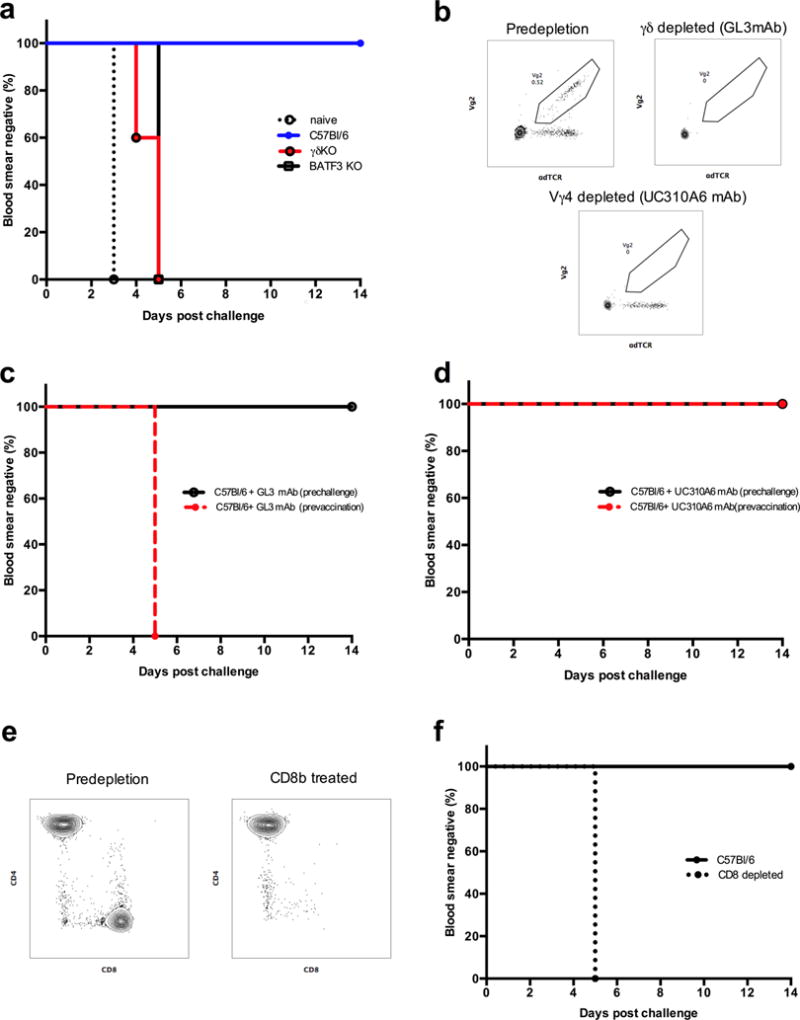
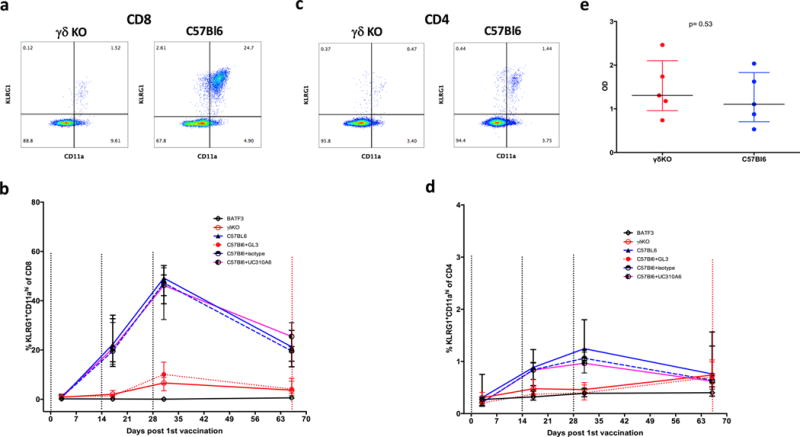
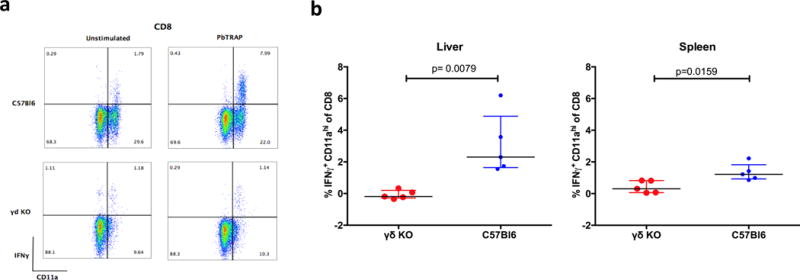
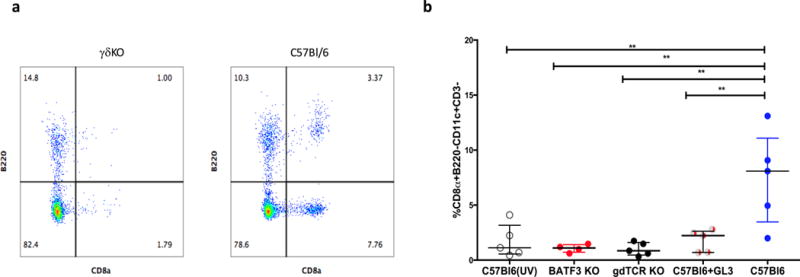
Whole-sporozoite vaccines confer sterilizing immunity to malaria-naive individuals by unknown mechanisms. In the first PfSPZ Vaccine trial ever in a malaria-endemic population, Vδ2 γδ T cells were significantly elevated and Vγ9/Vδ2 transcripts ranked as the most upregulated in vaccinees who were protected from Plasmodium falciparum infection. In a mouse model, absence of γδ T cells during vaccination impaired protective CD8 T cell responses and ablated sterile protection. γδ T cells were not required for circumsporozoite protein-specific Ab responses, and γδ T cell depletion before infectious challenge did not ablate protection. γδ T cells alone were insufficient to induce protection and required the presence of CD8α+ dendritic cells. In the absence of γδ T cells, CD8α+ dendritic cells did not accumulate in the livers of vaccinated mice. Altogether, our results show that γδ T cells were essential for the induction of sterile immunity during whole-organism vaccination.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures
Comment in
-
Comment on "γδ T Cells Are Required for the Induction of Sterile Immunity during Irradiated Sporozoite Vaccinations".J Immunol. 2018 Mar 1;200(5):1533. doi: 10.4049/jimmunol.1701743. J Immunol. 2018. PMID: 29463687 No abstract available.
-
Response to Comment on "γδ T Cells Are Required for the Induction of Sterile Immunity during Irradiated Sporozoite Vaccinations".J Immunol. 2018 Mar 1;200(5):1533-1534. doi: 10.4049/jimmunol.1701799. J Immunol. 2018. PMID: 29463688 Free PMC article. No abstract available.
References
-
- Epstein JE, Tewari K, Lyke KE, Sim BK, Billingsley PF, Laurens MB, Gunasekera A, Chakravarty S, James ER, Sedegah M, Richman A, Velmurugan S, Reyes S, Li M, Tucker K, Ahumada A, Ruben AJ, Li T, Stafford R, Eappen AG, Tamminga C, Bennett JW, Ockenhouse CF, Murphy JR, Komisar J, Thomas N, Loyevsky M, Birkett A, Plowe CV, Loucq C, Edelman R, Richie TL, Seder RA, Hoffman SL. Live attenuated malaria vaccine designed to protect through hepatic CD8(+) T cell immunity. Science. 2011;334:475–480. - PubMed
-
- Seder RA, Chang LJ, Enama ME, Zephir KL, Sarwar UN, Gordon IJ, Holman LA, James ER, Billingsley PF, Gunasekera A, Richman A, Chakravarty S, Manoj A, Velmurugan S, Li M, Ruben AJ, Li T, Eappen AG, Stafford RE, Plummer SH, Hendel CS, Novik L, Costner PJ, Mendoza FH, Saunders JG, Nason MC, Richardson JH, Murphy J, Davidson SA, Richie TL, Sedegah M, Sutamihardja A, Fahle GA, Lyke KE, Laurens MB, Roederer M, Tewari K, Epstein JE, Sim BK, Ledgerwood JE, Graham BS, Hoffman SL, V. R. C. S. Team Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science. 2013;341:1359–1365. - PubMed
-
- Hoffman SL, Billingsley PF, James E, Richman A, Loyevsky M, Li T, Chakravarty S, Gunasekera A, Chattopadhyay R, Li M, Stafford R, Ahumada A, Epstein JE, Sedegah M, Reyes S, Richie TL, Lyke KE, Edelman R, Laurens MB, Plowe CV, Sim BK. Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum malaria. Hum Vaccin. 2010;6:97–106. - PubMed
-
- Ishizuka AS, Lyke KE, DeZure A, Berry AA, Richie TL, Mendoza FH, Enama ME, Gordon IJ, Chang LJ, Sarwar UN, Zephir KL, Holman LA, James ER, Billingsley PF, Gunasekera A, Chakravarty S, Manoj A, Li M, Ruben AJ, Li T, Eappen AG, Stafford RE, CN K, Murshedkar T, DeCederfelt H, Plummer SH, Hendel CS, Novik L, Costner PJ, Saunders JG, Laurens MB, Plowe CV, Flynn B, Whalen WR, Todd JP, Noor J, Rao S, Sierra-Davidson K, Lynn GM, Epstein JE, Kemp MA, Fahle GA, Mikolajczak SA, Fishbaugher M, Sack BK, Kappe SH, Davidson SA, Garver LS, Bjorkstrom NK, Nason MC, Graham BS, Roederer M, Sim BK, Hoffman SL, Ledgerwood JE, Seder RA. Protection against malaria at 1year and immune correlates following PfSPZ vaccination. Nat Med 2016 - PMC - PubMed
-
- Holtmeier W, Kabelitz D. gammadelta T cells link innate and adaptive immune responses. Chem Immunol Allergy. 2005;86:151–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
