

Review
doi: 10.1021/acschembio.7b00760.
Epub 2017 Nov 14.
Repair of a Site-Specific DNA Cleavage: Old-School Lessons for Cas9-Mediated Gene Editing
Affiliations
- PMID: 29083855
- PMCID: PMC5835394
- DOI: 10.1021/acschembio.7b00760
Item in Clipboard
Review
Repair of a Site-Specific DNA Cleavage: Old-School Lessons for Cas9-Mediated Gene Editing
ACS Chem Biol.
.
Abstract
CRISPR/Cas9-mediated gene editing may involve nonhomologous end-joining to create various insertion/deletions (indels) or may employ homologous recombination to modify precisely the target DNA sequence. Our understanding of these processes has been guided by earlier studies using other site-specific endonucleases, both in model organisms such as budding yeast and in mammalian cells. We briefly review what has been gleaned from such studies using the HO and I-SceI endonucleases and how these findings guide current gene editing strategies.
Figures
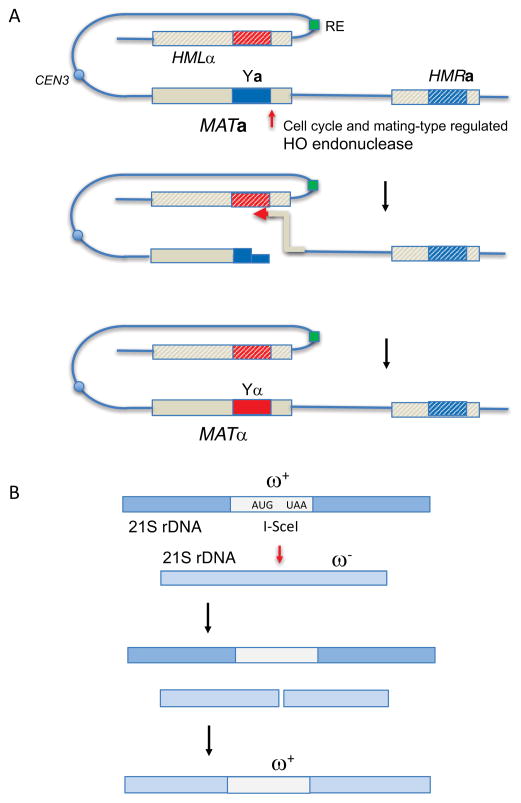
HO and I-SceI-mediated recombination in budding yeast. A. Mating-type (MAT) gene switching. Prior to S phase, budding yeast express HO endonuclease, resulting in the cleavage of either MATa or MATα on chromosome 3. The silent, heterochromatic structure (hatched lines) of the two donors, HMLα and HMRa, located 200 and 100 kb from MAT, respectively, are resistant to cleavage, but serve as efficient donors to repair the DSB. Donor preference is regulated by the cis-acting Recombination Enhancer (RE); in MATa cells, proteins bound to RE also bind near the DSB end and promotes use of HML. In MATα, RE is itself silenced, allowing HMRa to be the default donor. Repair of the DSB involves removal of the original mating type-specific Ya or Yα sequences and their replacement with the opposite mating type. DSB repair by gene conversion without an accompanying exchange (see Fig. 2A) is the predominant mechanism of repair. B. Intron homing of the omega ω+ intron within the mitochondrial 21S ribosomal DNA. When an ω+ budding yeast mates with an ω− cell of opposite mating type, I-SceI endonuclease, encoded within the intron, cleaves its target site in the ω− locus and promotes repair by gene conversion. Details of the molecular events that lead to repair have not been established in detail. As there are often multiple copies of each mitochondrial genome, the global replacement of ω− by ω+ is the first example of gene drive.
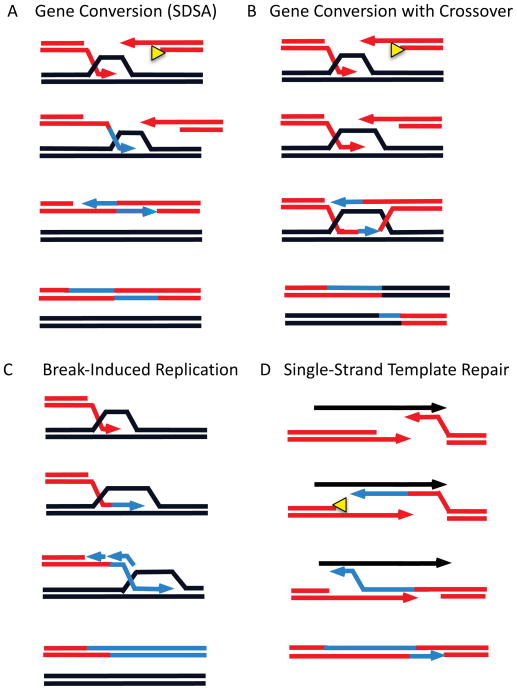
Mechanisms to repair a double-strand break. A. Gene conversion by synthesis-dependent strand annealing (SDSA) allows repair by patching up a DSB without chromosomal rearrangement. Key steps include 5′ to 3′ resection of DSB ends, recruitment of Rad51 recombinase as a filament on the 3′-ended single-stranded DNA (ssDNA), a homology search culminating in the formation of a displacement loop (D-loop) by base-pairing between the invading strand and the complementary homologous strand in the donor locus, initiation of new DNA synthesis, displacement of the newly synthesized DNA to anneal with the second end of the DSB (second end-capture), and completion of repair by second-end DNA synthesis and ligation of the strands. B. Gene conversion via a double-Holliday junction (dHJ) intermediate follows the same initial steps but an extended D-loop is annealed by the second end. New DNA synthesis and ligation lead to a dHJ structure that can be “dissolved” to a non-crossover outcome or resolved by structure-specific nucleases to produce a crossover outcome. C. Break-induced replication (BIR) begins similarly to SDSA, but new DNA synthesis requires all of the normal replication apparatus plus at least two proteins not needed for gene conversion or normal DNA replication. Although both leading- and lagging-strand synthesis must occur, they are not coordinated as in normal replication and all the newly copied DNA is conservatively inherited at the initially broken end. D. In single-strand template repair (SSTR), only one resected end of the DSB can pair with the ssDNA template and initiate new DNA synthesis, but there is apparently no need for Rad51 and strand invasion to accomplish this step. The newly extended DNA is apparently displaced from the template and can anneal with the second end – analogous to second-end capture – and second end synthesis can complete the repair of the DSB.
References
-
- Stoddard BL. Homing endonuclease structure and function. Q Rev Biophys. 2005;38:49–95. - PubMed
-
- Lee CS, Haber JE. Mating-type Gene Switching in Saccharomyces cerevisiae. Microbiol Spectr. 2015;3 MDNA3-0013-2014. - PubMed
-
- Haber JE. A Life Investigating Pathways That Repair Broken Chromosomes. Annu Rev Genet. 2016;50:1–28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
