

AtHKT1 drives adaptation of Arabidopsis thaliana to salinity by reducing floral sodium content
- PMID: 29084222
- PMCID: PMC5679648
- DOI: 10.1371/journal.pgen.1007086
AtHKT1 drives adaptation of Arabidopsis thaliana to salinity by reducing floral sodium content
Abstract
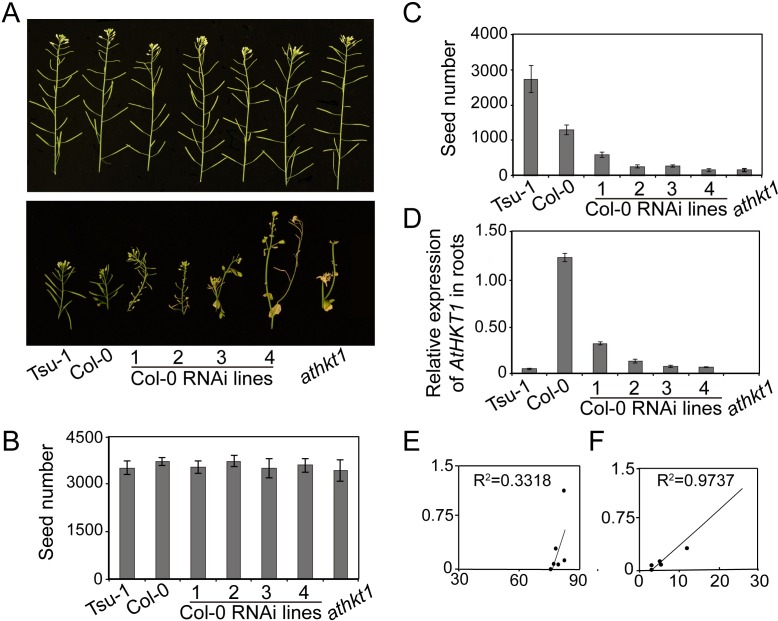
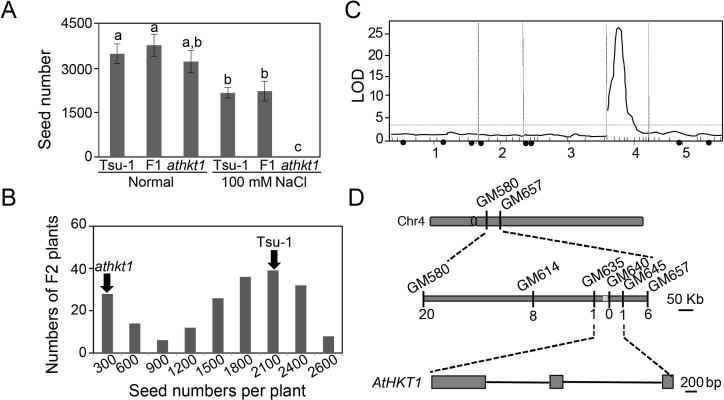
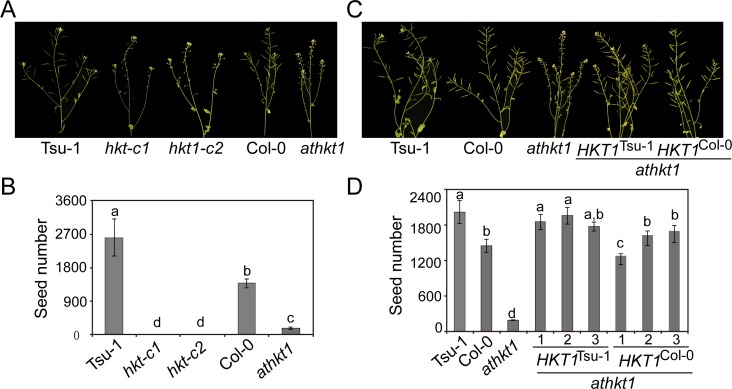
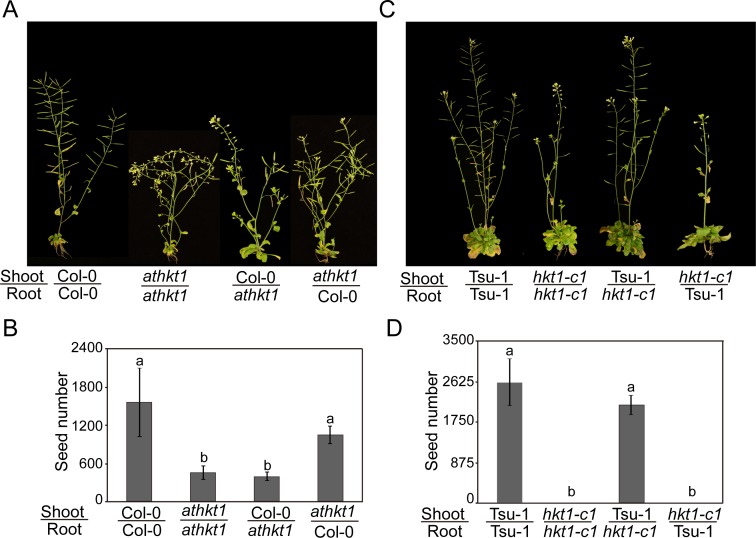
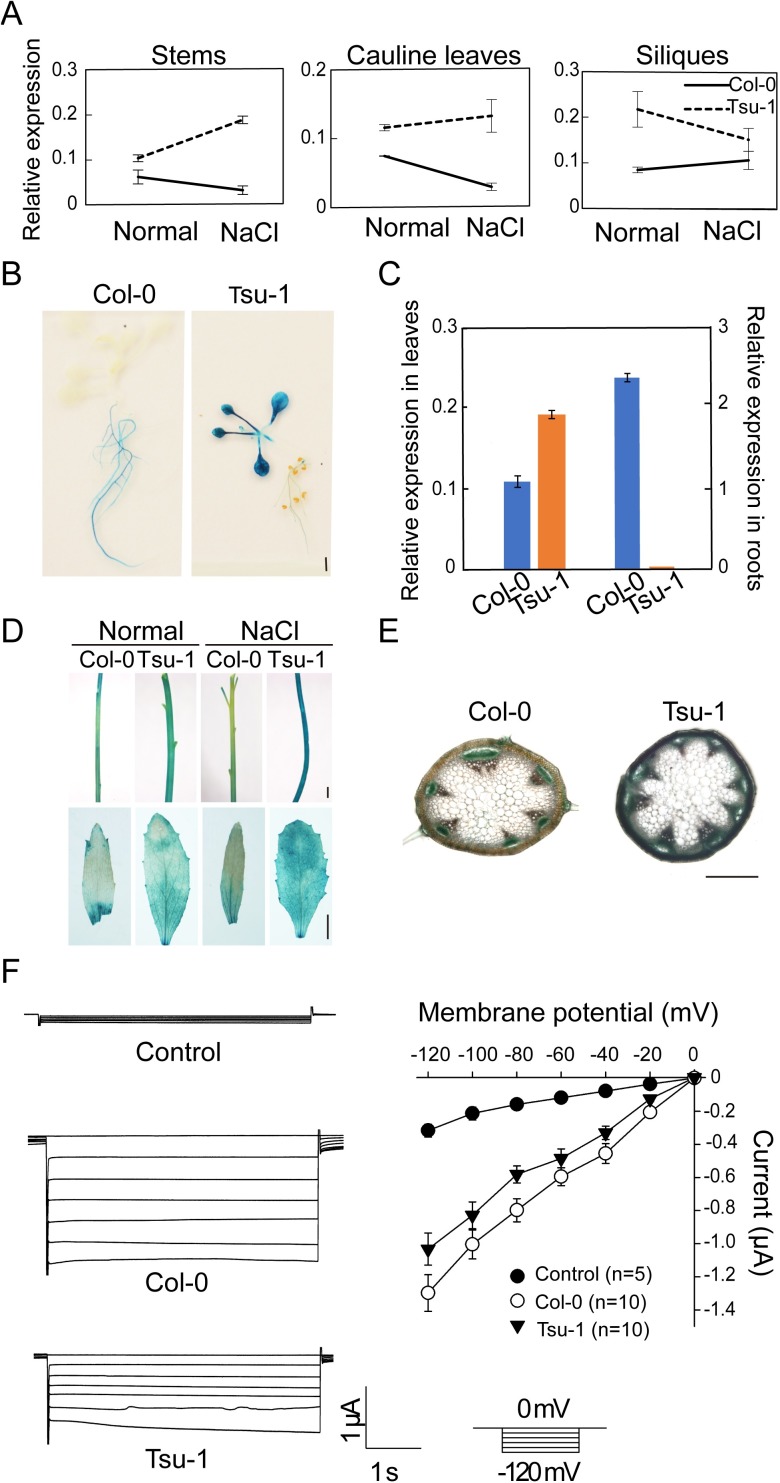
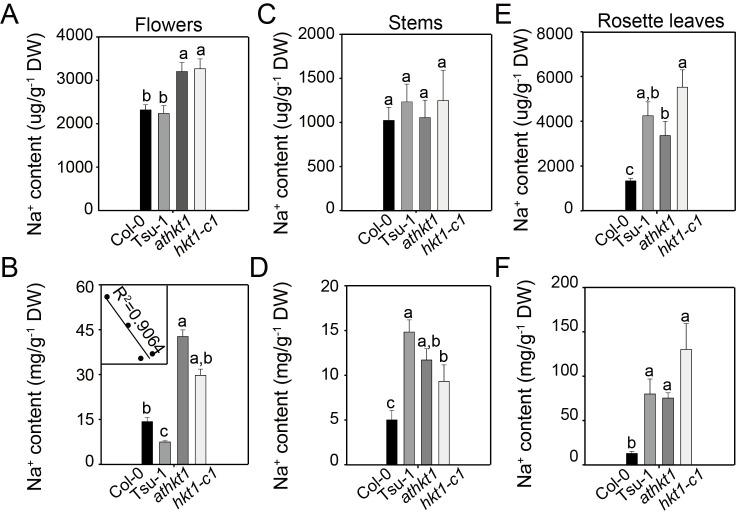
Arabidopsis thaliana high-affinity potassium transporter 1 (AtHKT1) limits the root-to-shoot sodium transportation and is believed to be essential for salt tolerance in A. thaliana. Nevertheless, natural accessions with 'weak allele' of AtHKT1, e.g. Tsu-1, are mainly distributed in saline areas and are more tolerant to salinity. These findings challenge the role of AtHKT1 in salt tolerance and call into question the involvement of AtHKT1 in salinity adaptation in A. thaliana. Here, we report that AtHKT1 indeed drives natural variation in the salt tolerance of A. thaliana and the coastal AtHKT1, so-called weak allele, is actually hyper-functional in reducing flowers sodium content upon salt stress. Our data showed that AtHKT1 positively contributes to saline adaptation in a linear manner. Forward and reverse genetics analysis established that the single AtHKT1 locus is responsible for the variation in the salinity adaptation between Col-0 and Tsu-1. Reciprocal grafting experiments revealed that shoot AtHKT1 determines the salt tolerance of Tsu-1, whereas root AtHKT1 primarily drives the salt tolerance of Col-0. Furthermore, evidence indicated that Tsu-1 AtHKT1 is highly expressed in stems and is more effective compared to Col-0 AtHKT1 at limiting sodium flow to the flowers. Such efficient retrieval of sodium to the reproductive organ endows Tsu-1 with stronger fertility compared to Col-0 upon salt stress, thus improving Tsu-1 adaptation to a coastal environment. To conclude, our data not only confirm the role of AtHKT1 in saline adaptation, but also sheds light on our understanding of the salt tolerance mechanisms in plants.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Ma Y, Dai X, Xu Y, Luo W, Zheng X, Zeng D, et al. COLD1 confers chilling tolerance in rice. Cell. 2015;160: 1209–1221. doi: 10.1016/j.cell.2015.01.046 - DOI - PubMed
-
- Li Y, Huang Y, Bergelson J, Nordborg M, Borevitz JO. Association mapping of local climate-sensitive quantitative trait loci in Arabidopsis thaliana. Proc Natl Acad Sci USA. 2010;107: 21199–21204. doi: 10.1073/pnas.1007431107 - DOI - PMC - PubMed
-
- Lu S, Zhao X, Hu Y, Liu S, Nan H, Li X, et al. Natural variation at the soybean J locus improves adaptation to the tropics and enhances yield. Nat Genet. 2017;49: 773–779. doi: 10.1038/ng.3819 - DOI - PubMed
-
- Todesco M, Balasubramanian S, Hu TT, Traw MB, Horton M, Epple P, et al. Natural allelic variation underlying a major fitness trade-off in Arabidopsis thaliana. Nature. 2010;465: 632–636. doi: 10.1038/nature09083 - DOI - PMC - PubMed
-
- Chao D-Y, Dilkes B, Luo H, Douglas A, Yakubova E, Lahner B, et al. Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis. Science. 2013;341: 658–659. doi: 10.1126/science.1240561 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
