

Hematopoietic Stem Cells: Normal Versus Malignant
- PMID: 29084438
- PMCID: PMC6207150
- DOI: 10.1089/ars.2017.7326
Hematopoietic Stem Cells: Normal Versus Malignant
Abstract
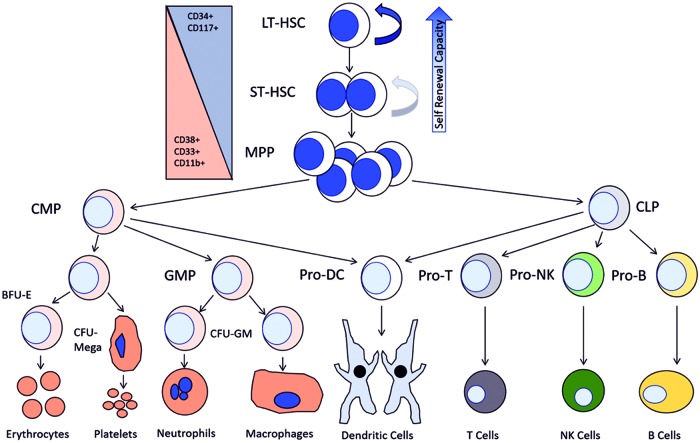
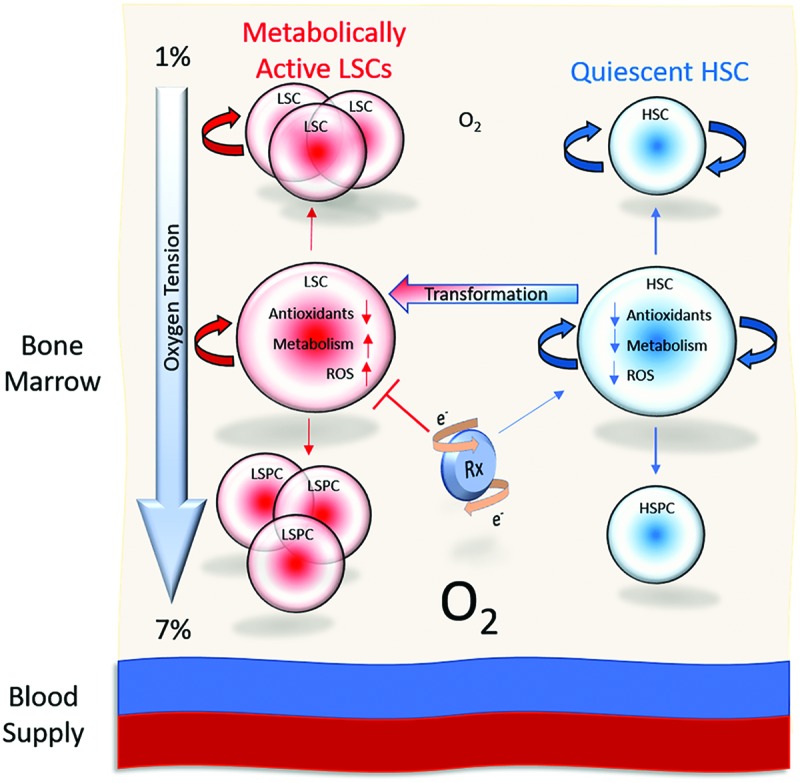
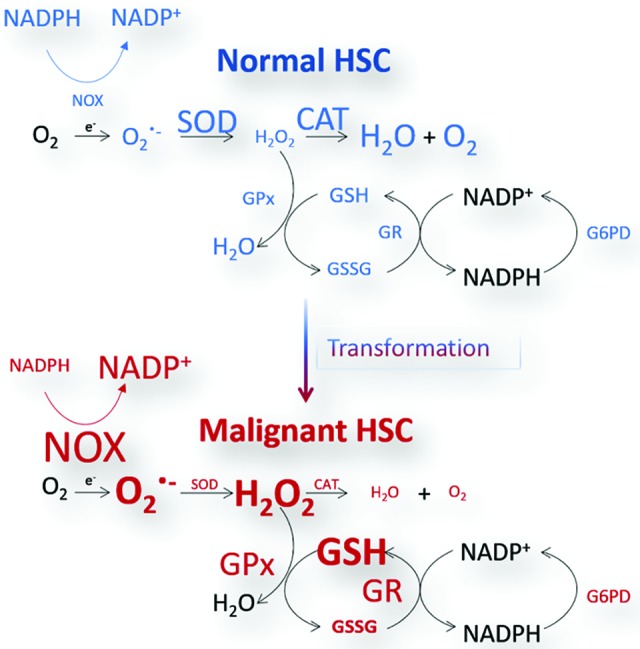
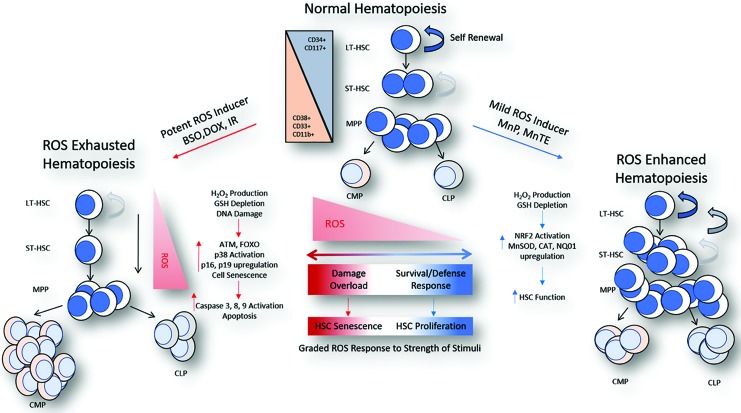
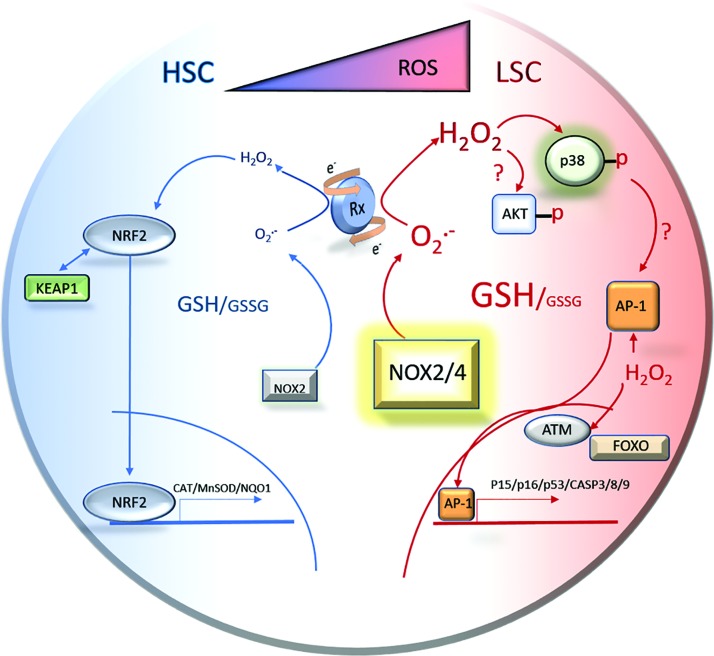
Significance: The long-term hematopoietic stem cell (LT-HSC) demonstrates characteristics of self-renewal and the ability to manage expansion of the hematopoietic compartment while maintaining the capacity for differentiation into hematopoietic stem/progenitor cell (HSPC) and terminal subpopulations. Deregulation of the HSPC redox environment results in loss of signaling that normally controls HSPC fate, leading to a loss of HSPC function and exhaustion. The characteristics of HSPC exhaustion via redox stress closely mirror phenotypic traits of hematopoietic malignancies and the leukemic stem cell (LSC). These facets elucidate the HSC/LSC redox environment as a druggable target and a growing area of cancer research. Recent Advances: Although myelosuppression and exhaustion of the hematopoietic niche are detrimental side effects of classical chemotherapies, new agents that modify the HSPC/LSC redox environment have demonstrated the potential for protection of normal HSPC function while inducing cytotoxicity within malignant populations.
Critical issues: New therapies must preserve, or only slightly disturb normal HSPC redox balance and function, while simultaneously altering the malignant cellular redox state. The cascade nature of redox damage makes this a critical and delicate line for the development of a redox-based therapeutic index.
Future directions: Recent evidence demonstrates the potential for redox-based therapies to impact metabolic and epigenetic factors that could contribute to initial LSC transformation. This is balanced by the development of therapies that protect HSPC function. This pushes toward therapies that may alter the HSC/LSC redox state but lead to initiation cell fate signaling lost in malignant transformation while protecting normal HSPC function. Antioxid. Redox Signal.
Keywords: HSC; LSC; hematopoiesis; redox-active compound; stem cell function.
Figures

References
-
- Adams GB. and Scadden DT. The hematopoietic stem cell in its place. Nat Immunol 7: 333–337, 2006 - PubMed
-
- Al-Hajj M, Becker MW, Wicha M, Weissman I, and Clarke MF. Therapeutic implications of cancer stem cells. Curr Opin Genet Dev 14: 43–47, 2004 - PubMed
-
- Allay JA, Spencer HT, Wilkinson SL, Belt JA, Blakley RL, and Sorrentino BP. Sensitization of hematopoietic stem and progenitor cells to trimetrexate using nucleoside transport inhibitors. Blood 90: 3546–3554, 1997 - PubMed
-
- Arai F, Hirao A, and Suda T. Regulation of hematopoietic stem cells by the niche. Trends Cardiovasc Med 15: 75–79, 2005 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
