

Gene Regulation and Survival under Hypoxia Requires Starch Availability and Metabolism
- PMID: 29084901
- PMCID: PMC5813553
- DOI: 10.1104/pp.17.01002
Gene Regulation and Survival under Hypoxia Requires Starch Availability and Metabolism
Abstract
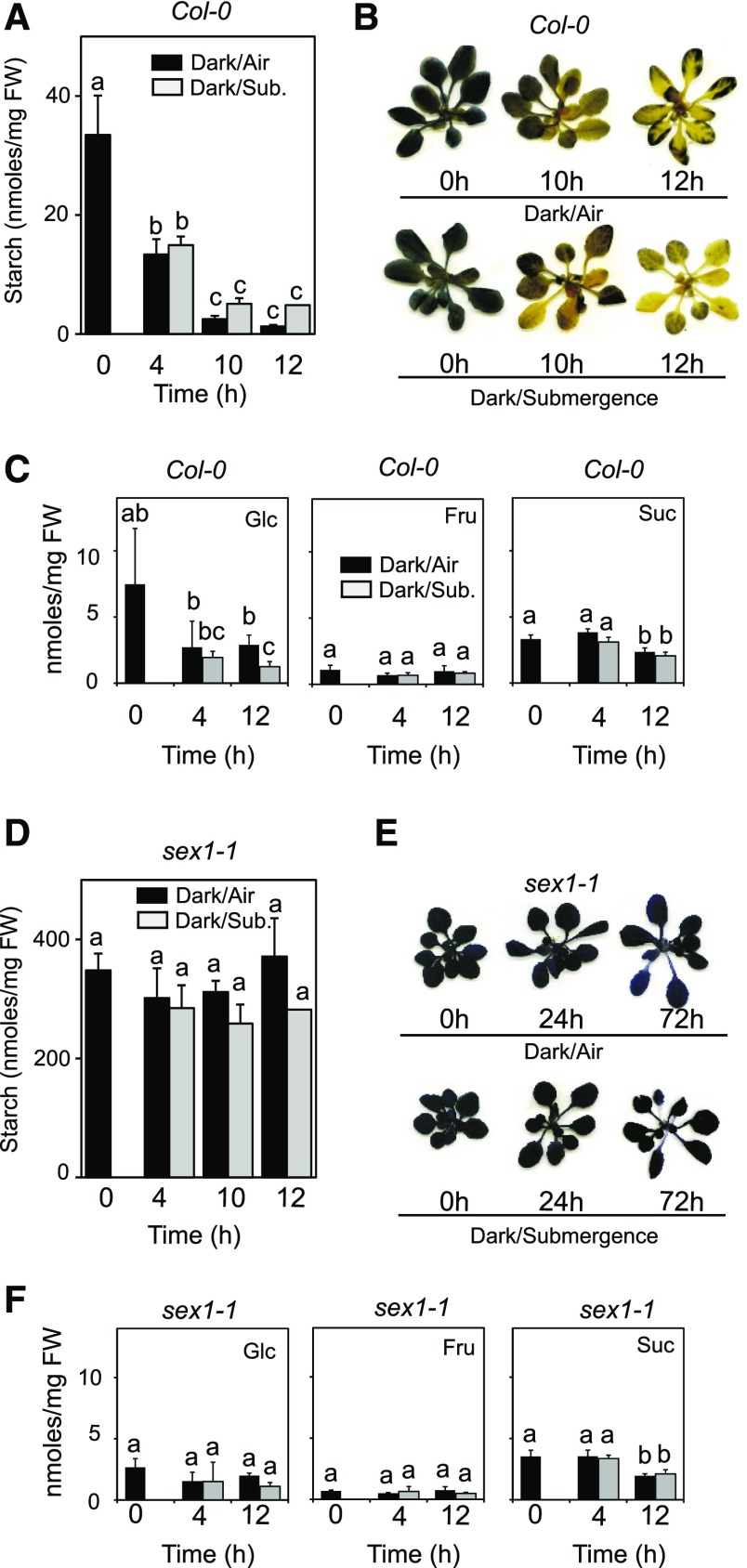
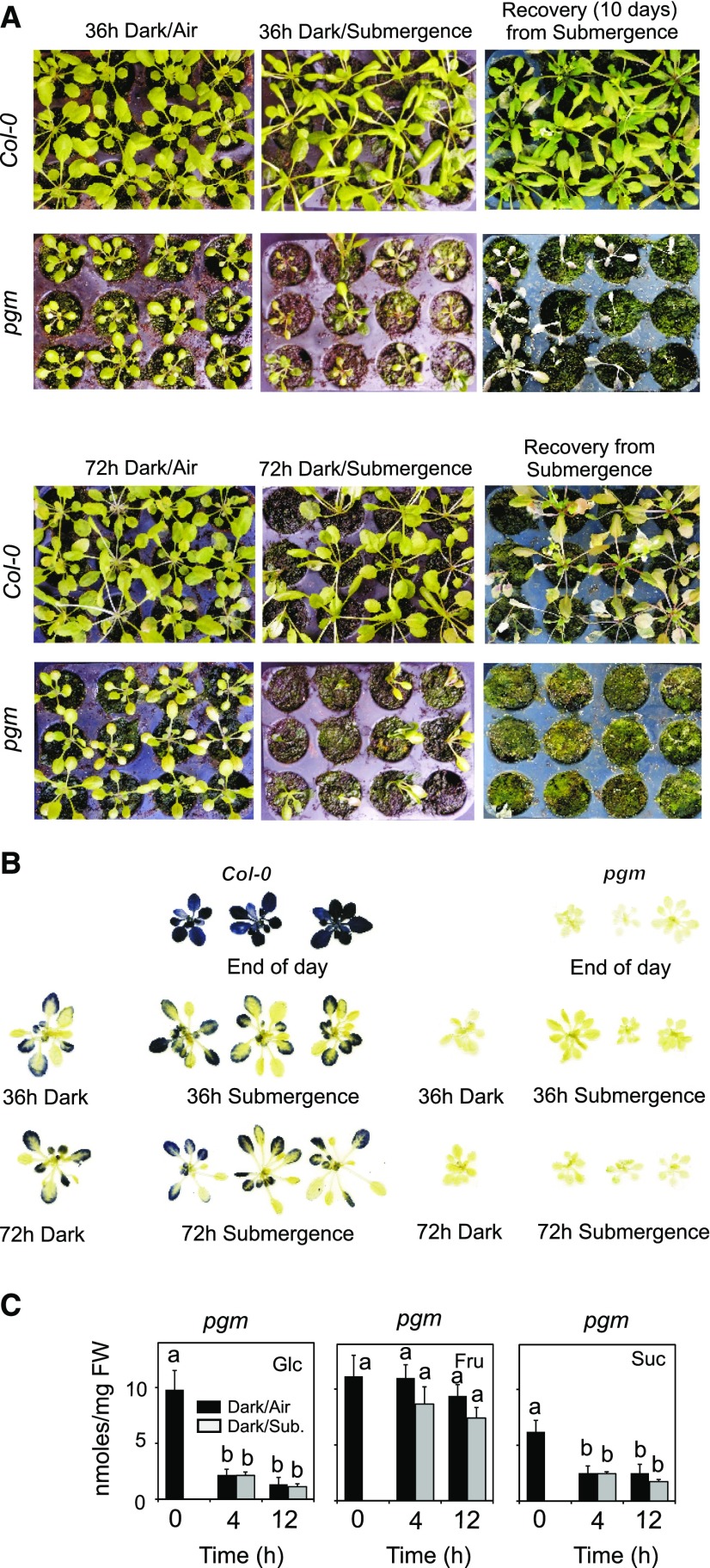
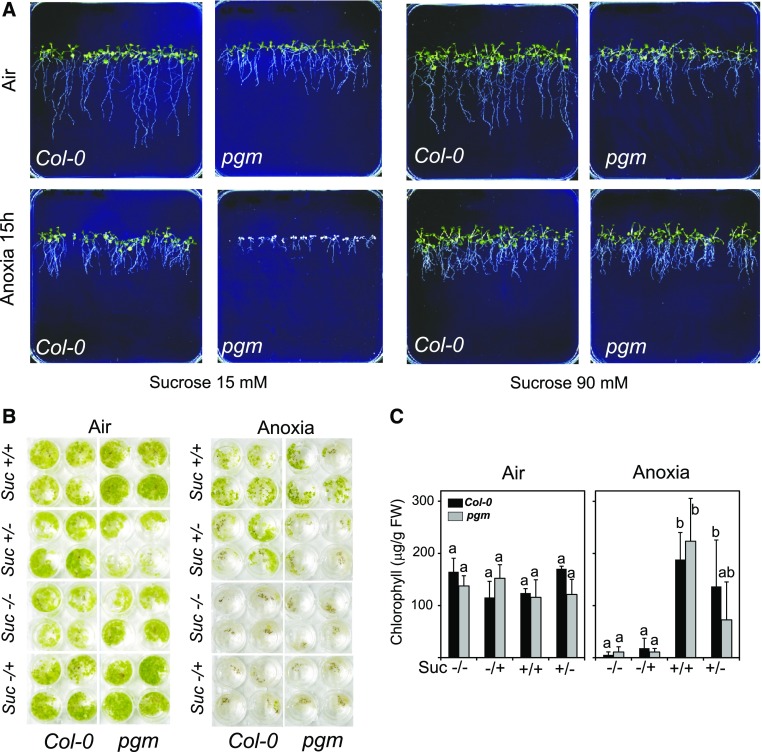
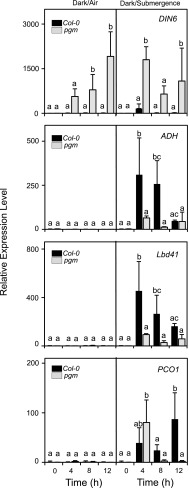
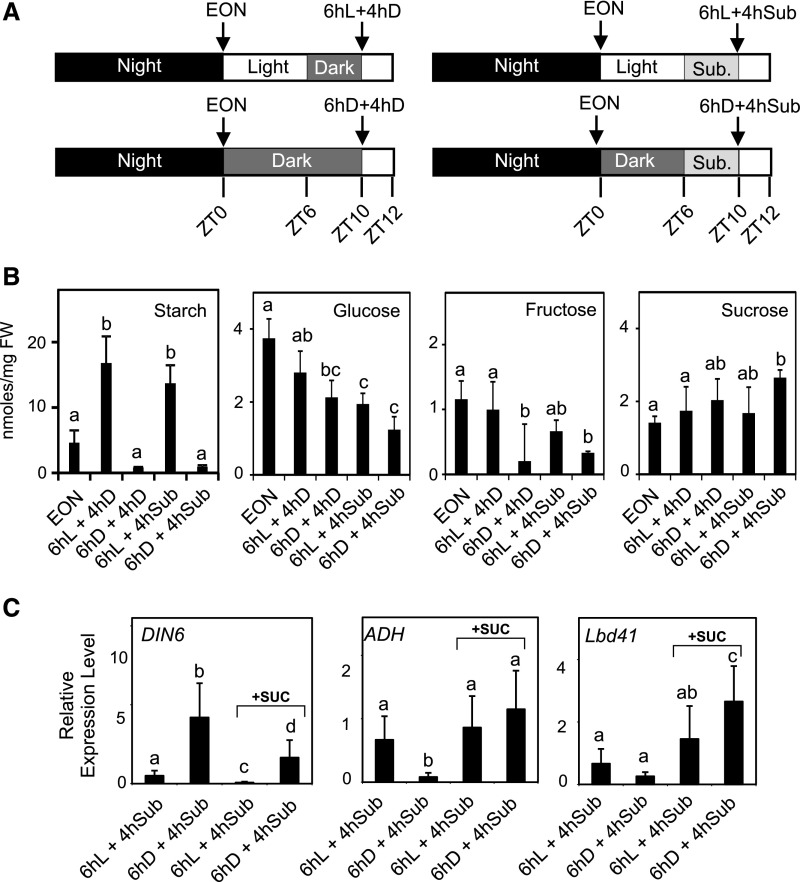
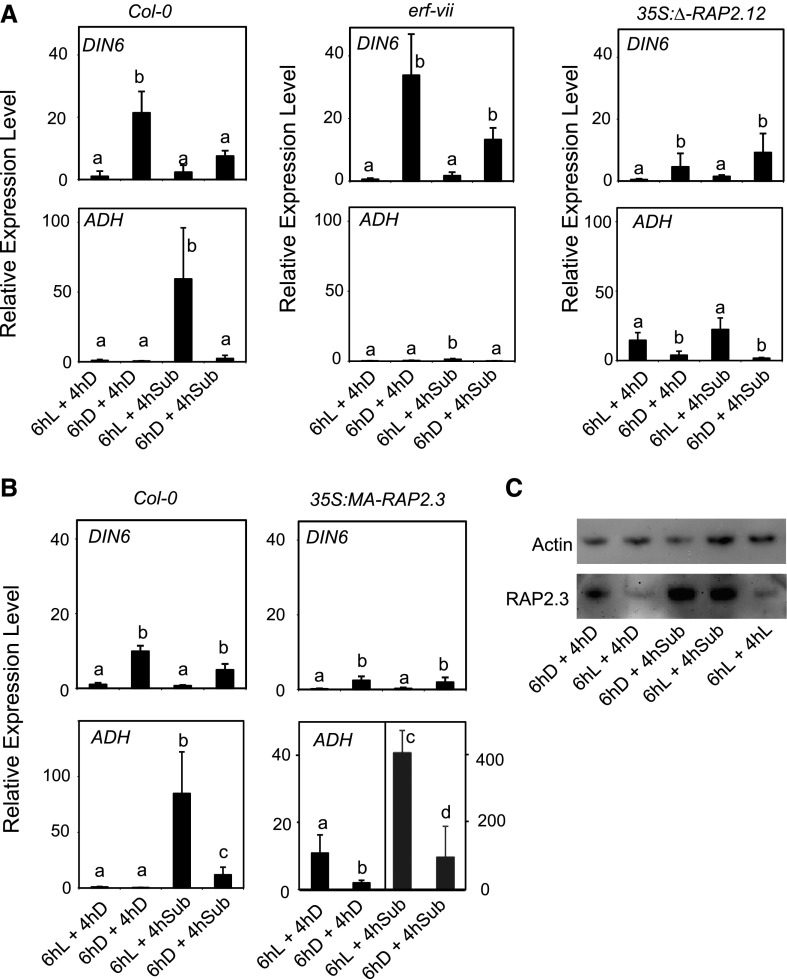
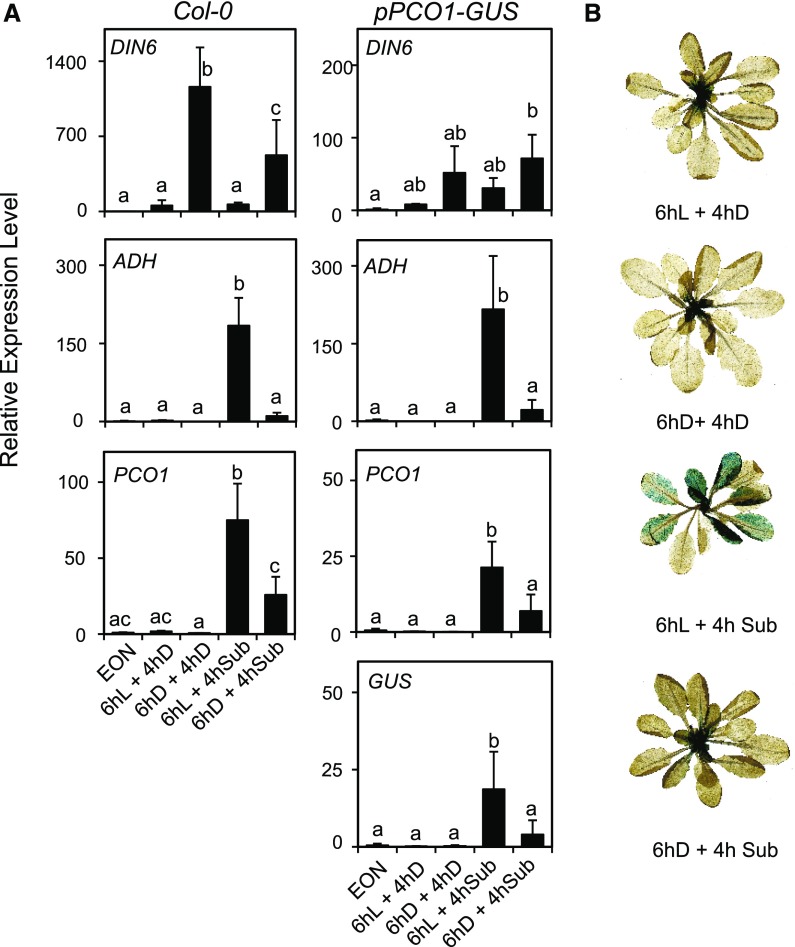
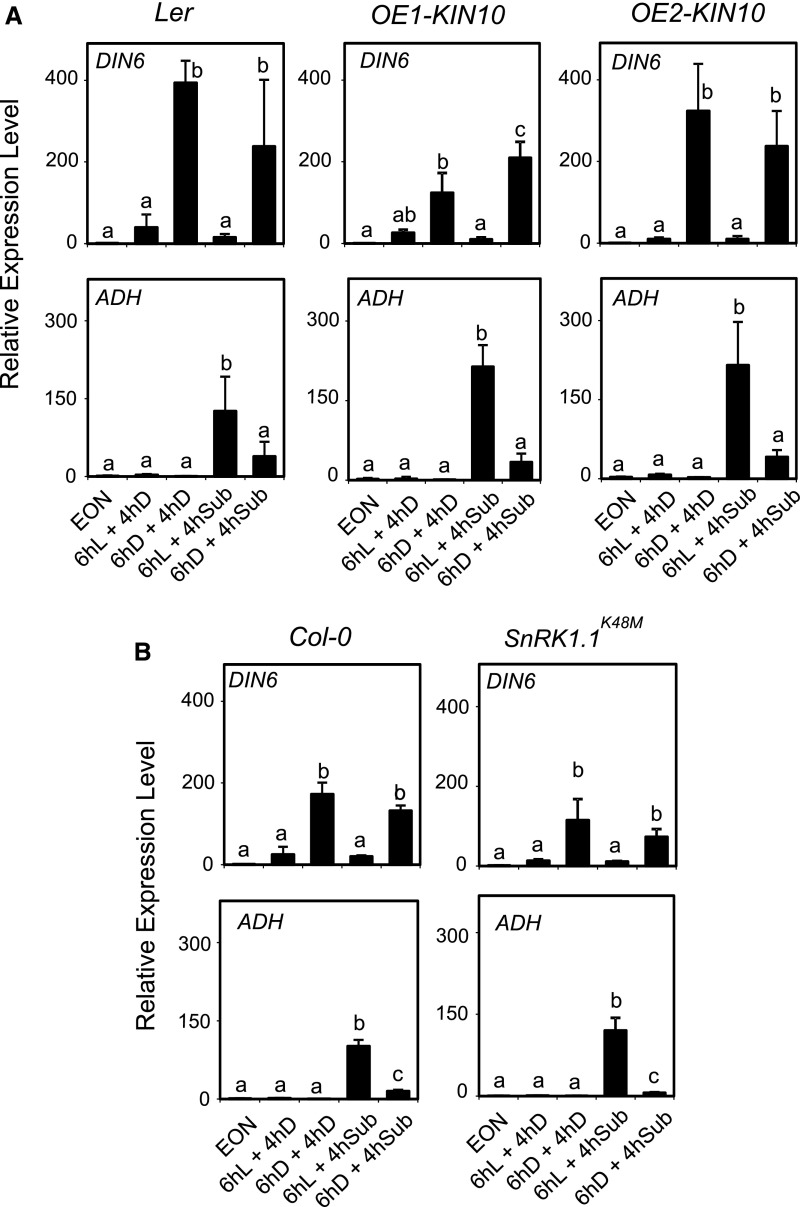
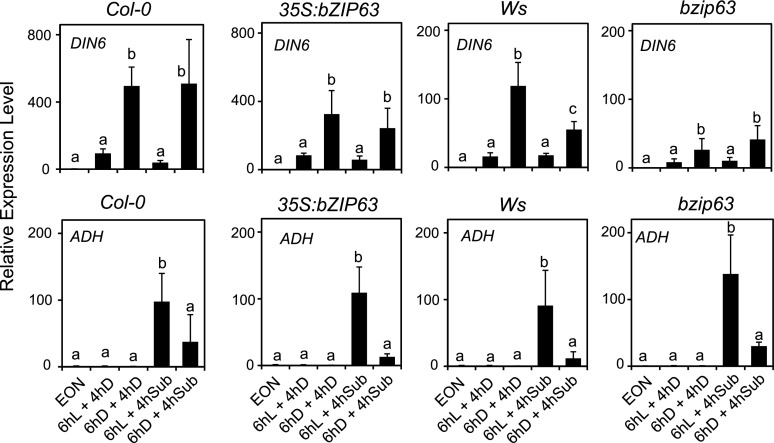
Plants respond to hypoxia, often caused by submergence, by expressing a specific set of genes that contribute to acclimation to this unfavorable environmental condition. Genes induced by low oxygen include those encoding enzymes for carbohydrate metabolism and fermentation, pathways that are required for survival. Sugar availability is therefore of crucial importance for energy production under hypoxia. Here, we show that Arabidopsis (Arabidopsis thaliana) plants require starch for surviving submergence as well as for ensuring the rapid induction of genes encoding enzymes required for anaerobic metabolism. The starchless pgm mutant is highly susceptible to submergence and also fails to induce anaerobic genes at the level of the wild type. Treating wild-type plants under conditions inducing sugar starvation results in a weak induction of alcohol dehydrogenase and other anaerobic genes. Induction of gene expression under hypoxia requires transcription factors belonging to group VII ethylene response factors (ERF-VII) that, together with plant Cys oxidases, act as an oxygen-sensing mechanism. We show that repression of this pathway by sugar starvation occurs downstream of the hypoxia-dependent stabilization of ERF-VII proteins and independently of the energy sensor protein kinases SnRK1.1 (SNF1-related kinase 1.1).
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Baena-González E, Rolland F, Thevelein JM, Sheen J (2007) A central integrator of transcription networks in plant stress and energy signalling. Nature 448: 938–942 - PubMed
-
- Bailey-Serres J, Voesenek LACJ (2008) Flooding stress: acclimations and genetic diversity. Annu Rev Plant Biol 59: 313–339 - PubMed
-
- Bailey-Serres J, Fukao T, Gibbs DJ, Holdsworth MJ, Lee SC, Licausi F, Perata P, Voesenek LACJ, van Dongen JT (2012) Making sense of low oxygen sensing. Trends Plant Sci 17: 129–138 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
