

Relevance of iPSC-derived human PGC-like cells at the surface of embryoid bodies to prechemotaxis migrating PGCs
- PMID: 29087313
- PMCID: PMC5699045
- DOI: 10.1073/pnas.1707779114
Relevance of iPSC-derived human PGC-like cells at the surface of embryoid bodies to prechemotaxis migrating PGCs
Abstract
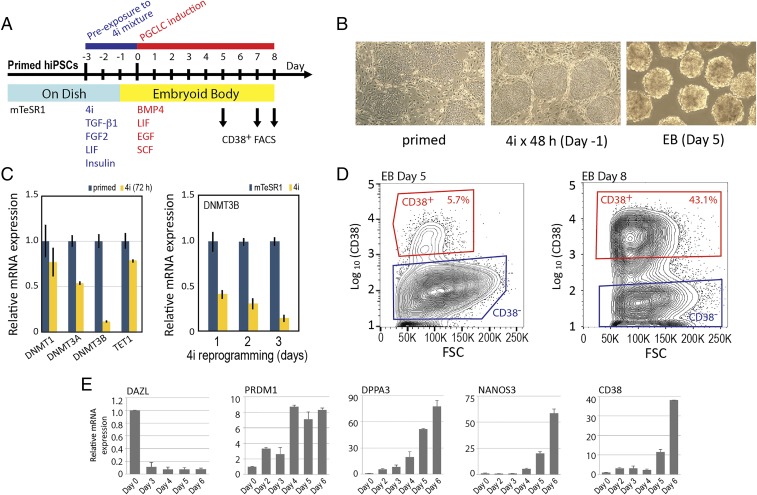
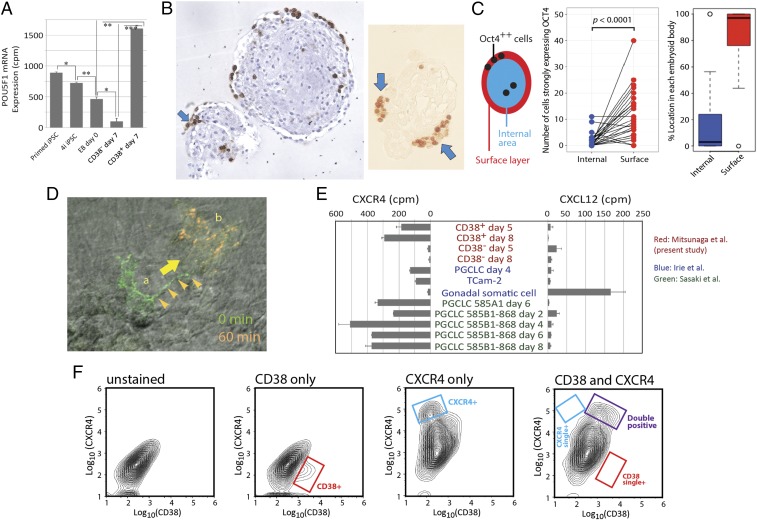
Pluripotent stem cell-derived human primordial germ cell-like cells (hPGCLCs) provide important opportunities to study primordial germ cells (PGCs). We robustly produced CD38+ hPGCLCs [∼43% of FACS-sorted embryoid body (EB) cells] from primed-state induced pluripotent stem cells (iPSCs) after a 72-hour transient incubation in the four chemical inhibitors (4i)-naïve reprogramming medium and showed transcriptional consistency of our hPGCLCs with hPGCLCs generated in previous studies using various and distinct protocols. Both CD38+ hPGCLCs and CD38- EB cells significantly expressed PRDM1 and TFAP2C, although PRDM1 mRNA in CD38- cells lacked the 3'-UTR harboring miRNA binding sites regulating mRNA stability. Genes up-regulated in hPGCLCs were enriched for cell migration genes, and their promoters were enriched for the binding motifs of TFAP2 (which was identified in promoters of T, NANOS3, and SOX17) and the RREB-1 cell adhesion regulator. In EBs, hPGCLCs were identified exclusively in the outermost surface monolayer as dispersed cells or cell aggregates with strong and specific expression of POU5F1/OCT4 protein. Time-lapse live cell imaging revealed active migration of hPGCLCs on Matrigel. Whereas all hPGCLCs strongly expressed the CXCR4 chemotaxis receptor, its ligand CXCL12/SDF1 was not significantly expressed in the whole EBs. Exposure of hPGCLCs to CXCL12/SDF1 induced cell migration genes and antiapoptosis genes. Thus, our study shows that transcriptionally consistent hPGCLCs can be readily produced from hiPSCs after transition of their pluripotency from the primed state using various methods and that hPGCLCs resemble the early-stage PGCs randomly migrating in the midline region of human embryos before initiation of the CXCL12/SDF1-guided chemotaxis.
Keywords: PRDM1; TFAP2C; embryoid body; induced pluripotent stem cells; primordial germ cells.
Copyright © 2017 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Weinberger L, Ayyash M, Novershtern N, Hanna JH. Dynamic stem cell states: Naive to primed pluripotency in rodents and humans. Nat Rev Mol Cell Biol. 2016;17:155–169. - PubMed
-
- Gafni O, et al. Derivation of novel human ground state naive pluripotent stem cells. Nature. 2013;504:282–286. - PubMed
-
- Magnúsdóttir E, Surani MA. How to make a primordial germ cell. Development. 2014;141:245–252. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
