

Review
doi: 10.1016/j.tibs.2017.10.001.
Epub 2017 Oct 28.
The Unsolved Problem of How Cells Sense Micron-Scale Curvature
Affiliations
- PMID: 29089160
- PMCID: PMC5705049
- DOI: 10.1016/j.tibs.2017.10.001
Item in Clipboard
Review
The Unsolved Problem of How Cells Sense Micron-Scale Curvature
Trends Biochem Sci.
2017 Dec.
Abstract
Membrane curvature is a fundamental feature of cells and their organelles. Much of what we know about how cells sense curved surfaces comes from studies examining nanometer-sized molecules on nanometer-scale curvatures. We are only just beginning to understand how cells recognize curved topologies at the micron scale. In this review, we provide the reader with an overview of our current understanding of how cells sense and respond to micron-scale membrane curvature.
Keywords: BAR domain; SpoVM; membrane curvature; septin.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
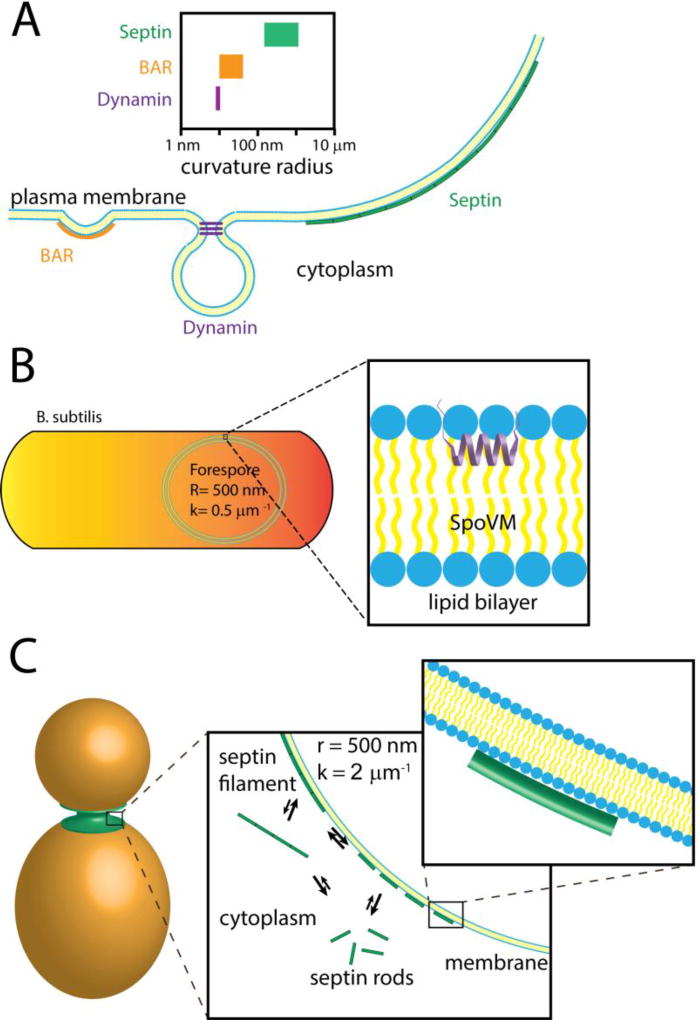
(A) Cells possess a variety of proteins that can bind and recognize curved biological membranes. Some of the best understood membrane curvature “sensors” are BAR domain proteins (left) and dynamin (center), which recognize steep, nanometer-scale curvatures. Comparatively, the septin cytoskeleton (right) recognizes much shallower curvatures at a micron-scale. The range of curvatures recognized by these proteins are depicted above. (B) During sporulation in the bacterium Bacillus subtilis SpoVM preferentially binds to the forespore membrane, the only site of positive curvature in the cell, which has a curvature of about 0.5 µm−1 (or a radius of 500 nm). The amphipathic helix of SpoVM is thought to insert into the lipid bilayer of the forespore membrane (inset). Across the length of SpoVM (~ 4 nm), the perceived membrane curvature is nearly flat. (C) After bud emergence in the budding yeast Saccharomyces cerevisiaie, septins are localized to the mother-bud neck, which is the only area of continuous positive curvature at the plasma membrane. Septins are thought to be in an equilibrium between soluble and assembled states at the plasma membrane (first inset). Septins may cooperatively associate with lipid supported bilayers on beads in vitro suggesting that septin polymerization into filaments stabilizes their association to the membrane. Across the length of septin rod (17–32 nm), the perceived membrane curvature is nearly flat (second inset).
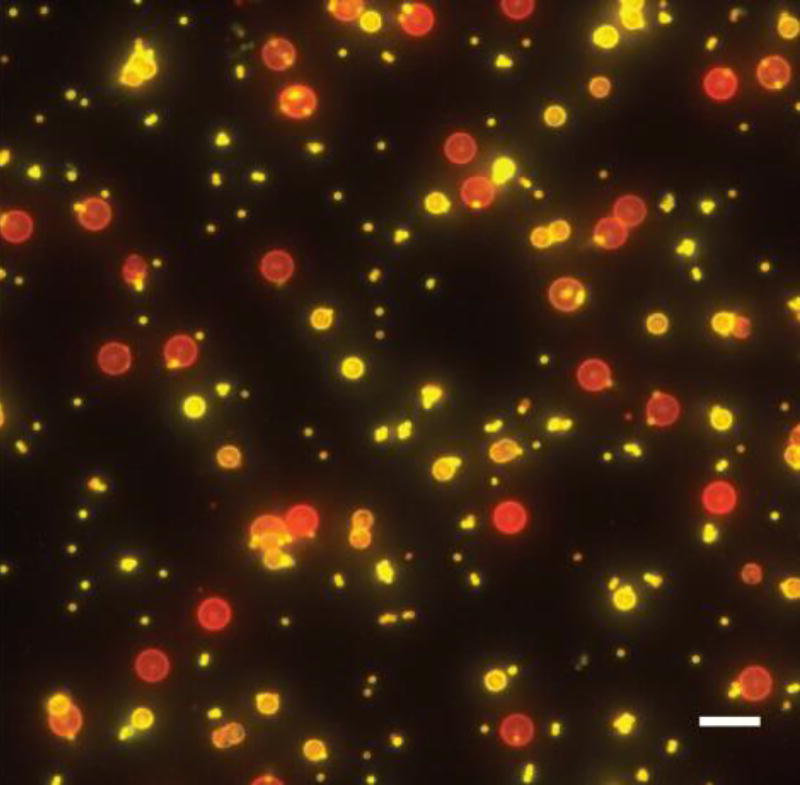
In vitro reconstitution of septins with lipid bilayer-coated beads (red) of various diameters shows septins enrichment on 1 µm and 3 µm beads over 5 µm beads (yellow). Scale bar, 10 µm
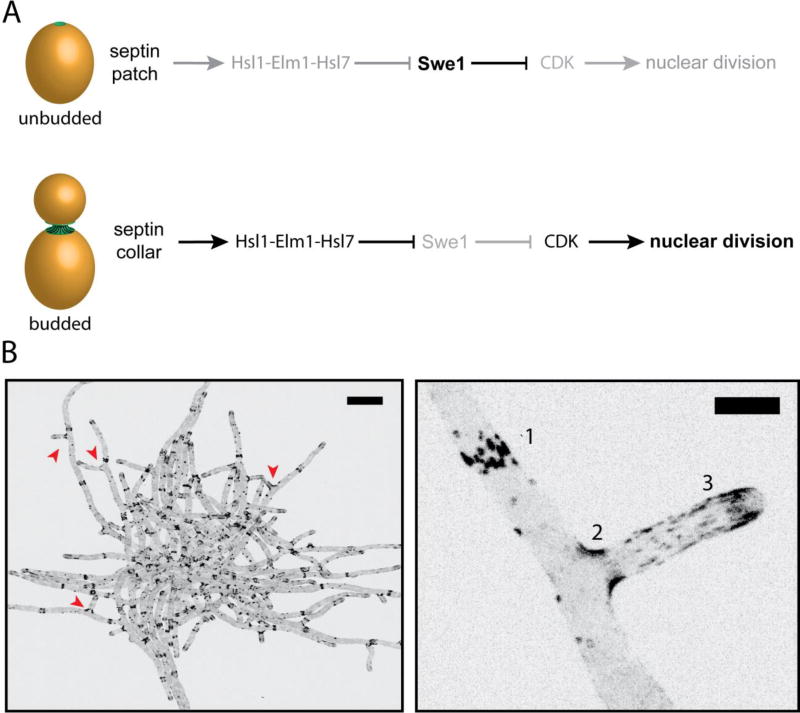
A. Morphogenesis checkpoint in S. cerevisiae: Hsl1-Elm1-Hsl7 are not recruited to the septin patch in unbudded cells, thereby allowing the CDK-inhibitor Swe1 levels to rise. As soon as the bud emerges, septin localization transitions to an hourglass shape coincident with the generation of micron-scale curvature. Swe1-regulators are recruited after this transition to initiate the degradation of Swe1 and promote progression through the cell cycle. B. An inverted, maximum z-stack projection of a A. gossypii cell expressing Cdc11-GFP. Scale bar, 20 µm. (Bottom right) Zoomed-in image of A. gossypii hyphae expressing Cdc11-GFP from above. Septins are organized into various higher-ordered structures in Ashbya: [1] Thick bars structures, [2] basal collars at branch points, and [3] thin, flexible filaments. Scale bar, 5 µm.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
