

The RNA exosome and RNA exosome-linked disease
- PMID: 29093021
- PMCID: PMC5769741
- DOI: 10.1261/rna.064626.117
The RNA exosome and RNA exosome-linked disease
Abstract
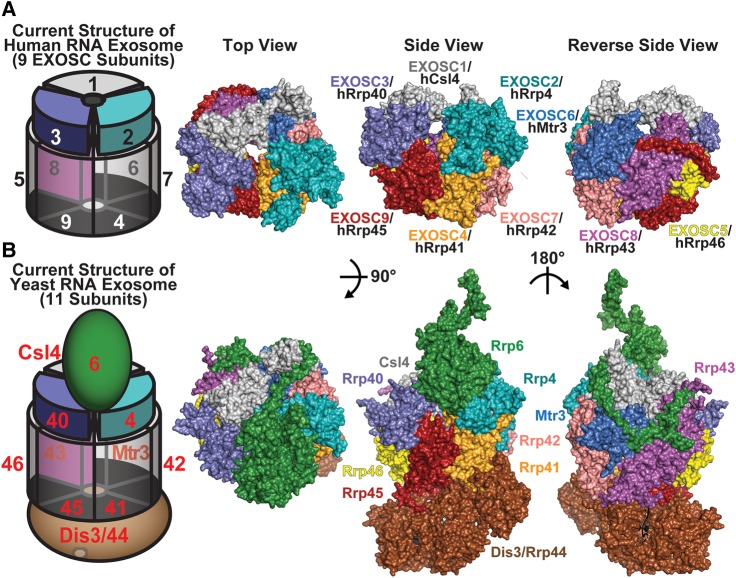
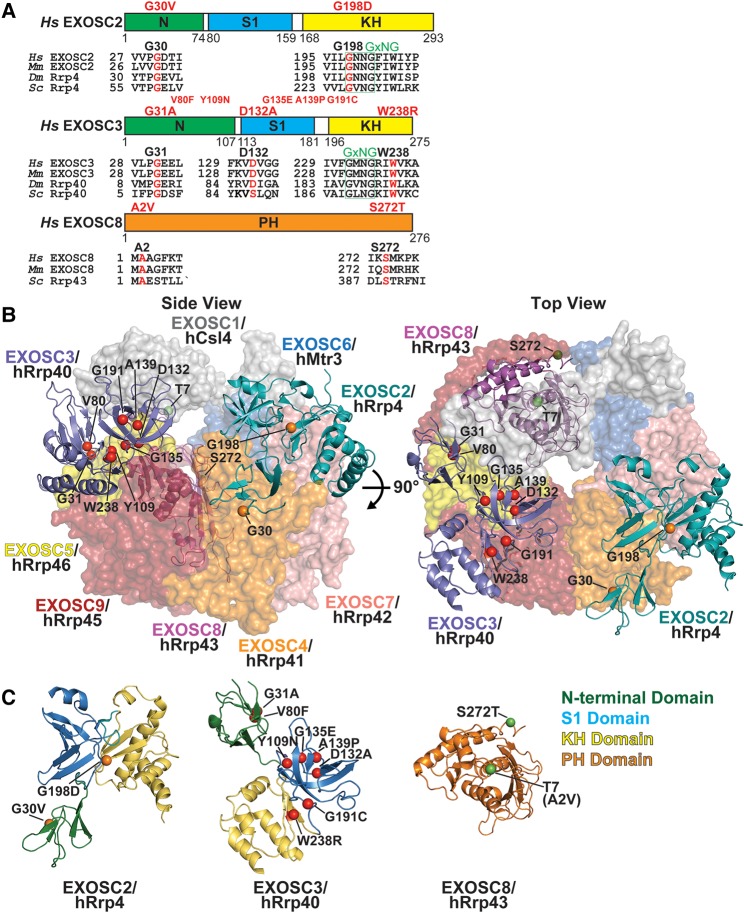
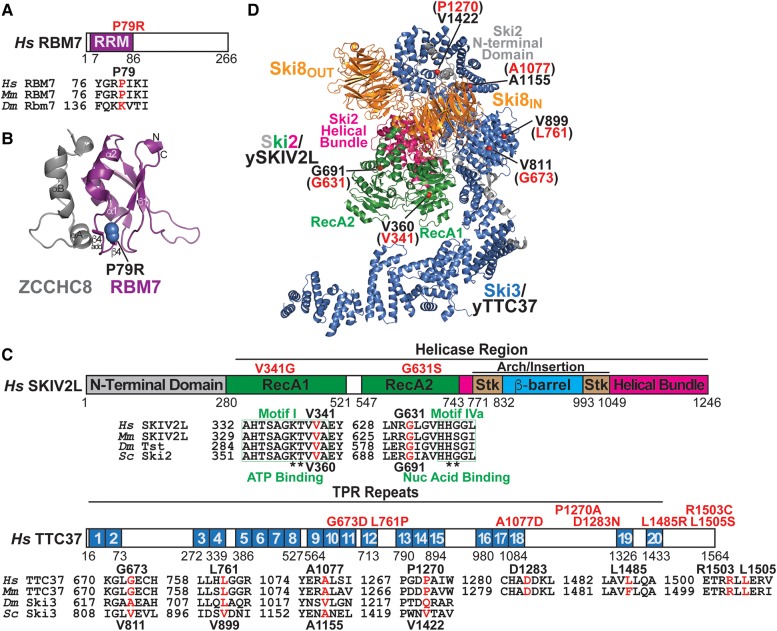
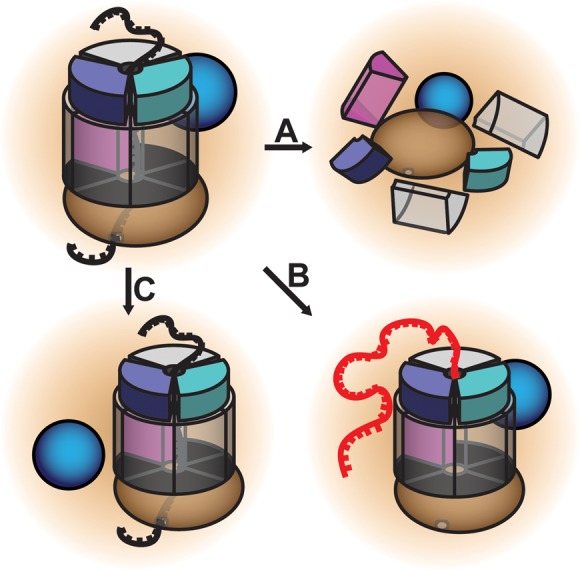
The RNA exosome is an evolutionarily conserved, ribonuclease complex that is critical for both processing and degradation of a variety of RNAs. Cofactors that associate with the RNA exosome likely dictate substrate specificity for this complex. Recently, mutations in genes encoding both structural subunits of the RNA exosome and its cofactors have been linked to human disease. Mutations in the RNA exosome genes EXOSC3 and EXOSC8 cause pontocerebellar hypoplasia type 1b (PCH1b) and type 1c (PCH1c), respectively, which are similar autosomal-recessive, neurodegenerative diseases. Mutations in the RNA exosome gene EXOSC2 cause a distinct syndrome with various tissue-specific phenotypes including retinitis pigmentosa and mild intellectual disability. Mutations in genes that encode RNA exosome cofactors also cause tissue-specific diseases with complex phenotypes. How mutations in these genes give rise to distinct, tissue-specific diseases is not clear. In this review, we discuss the role of the RNA exosome complex and its cofactors in human disease, consider the amino acid changes that have been implicated in disease, and speculate on the mechanisms by which exosome gene mutations could underlie dysfunction and disease.
Keywords: EXOSC2; EXOSC3; EXOSC8; RBM7; RNA exosome; RNA processing/degradation; Rrp4; Rrp40; Rrp43; SKIV2L; Ski2; Ski3; TTC37; intellectual disability; pontocerebellar hypoplasia type 1b; pontocerebellar hypoplasia type 1c; retinitis pigmentosa; spinal motor neuropathy; trichohepatoenteric syndrome.
© 2018 Morton et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous