

Lung-Infiltrating Foxp3+ Regulatory T Cells Are Quantitatively and Qualitatively Different during Eosinophilic and Neutrophilic Allergic Airway Inflammation but Essential To Control the Inflammation
- PMID: 29093062
- PMCID: PMC5716870
- DOI: 10.4049/jimmunol.1700211
Lung-Infiltrating Foxp3+ Regulatory T Cells Are Quantitatively and Qualitatively Different during Eosinophilic and Neutrophilic Allergic Airway Inflammation but Essential To Control the Inflammation
Abstract
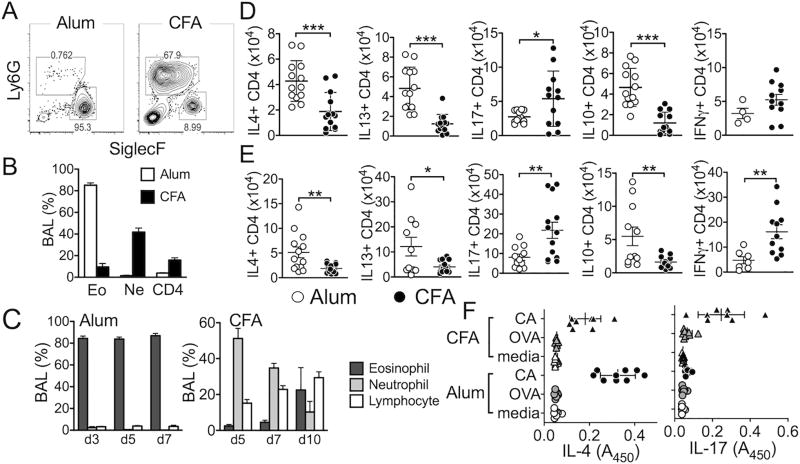
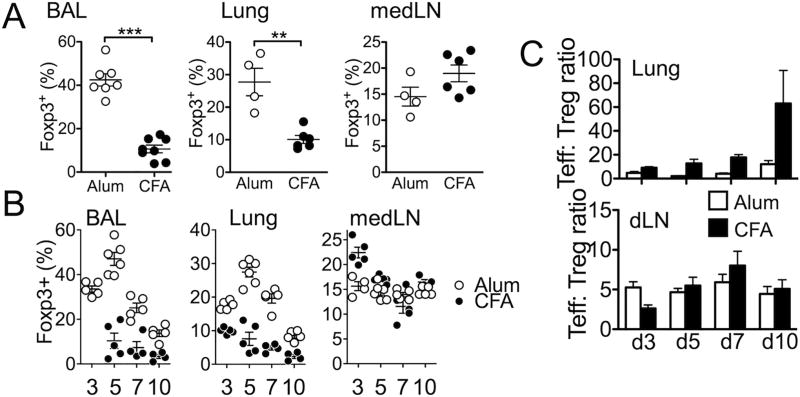
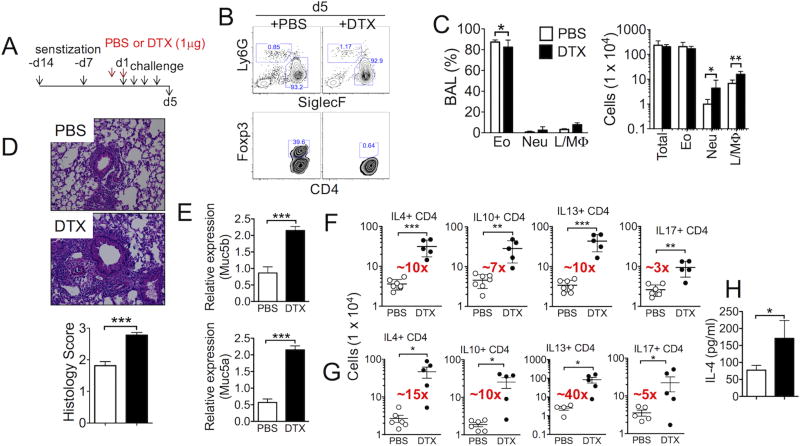
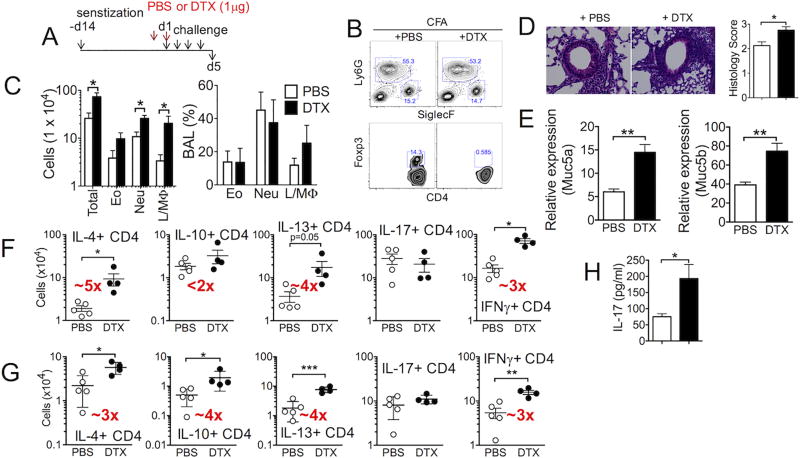
Understanding functions of Foxp3+ regulatory T cells (Tregs) during allergic airway inflammation remains incomplete. In this study, we report that, during cockroach Ag-induced allergic airway inflammation, Foxp3+ Tregs are rapidly mobilized into the inflamed lung tissues. However, the level of Treg accumulation in the lung was different depending on the type of inflammation. During eosinophilic airway inflammation, ∼30% of lung-infiltrating CD4 T cells express Foxp3, indicative of Tregs. On the contrary, only ∼10% of infiltrating CD4 T cells express Foxp3 during neutrophilic airway inflammation. Despite the different accumulation, the lung inflammation and inflammatory T cell responses were aggravated following Treg depletion, regardless of the type of inflammation, suggesting regulatory roles for Tregs. Interestingly, however, the extent to which inflammatory responses are aggravated by Treg depletion was significantly greater during eosinophilic airway inflammation. Indeed, lung-infiltrating Tregs exhibit phenotypic and functional features associated with potent suppression. Our results demonstrate that Tregs are essential regulators of inflammation, regardless of the type of inflammation, although the mechanisms used by Tregs to control inflammation may be shaped by environmental cues available to them.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures
References
-
- Tournoy KG, Provoost S, Van Hove C, Joos G. The role of immune tolerance in asthma pathogenesis. Curr Allergy Asthma Rep. 2006;6:437–443. - PubMed
-
- Holgate ST. Innate and adaptive immune responses in asthma. Nat Med. 2012;18:673–683. - PubMed
-
- Zhao Y, Yang J, Gao YD, Guo W. Th17 immunity in patients with allergic asthma. Int Arch Allergy Immunol. 2010;151:297–307. - PubMed
-
- Cosmi L, Liotta F, Maggi E, Romagnani S, Annunziato F. Th17 cells: new players in asthma pathogenesis. Allergy. 2011;66:989–998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
