

An Update on the Intracellular and Intercellular Trafficking of Carmoviruses
- PMID: 29093729
- PMCID: PMC5651262
- DOI: 10.3389/fpls.2017.01801
An Update on the Intracellular and Intercellular Trafficking of Carmoviruses
Abstract
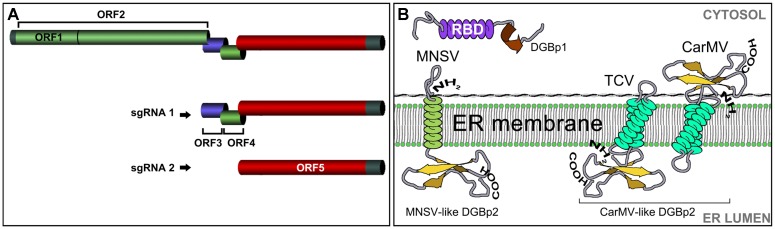
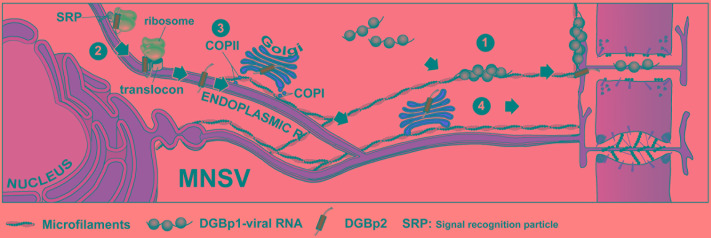
Despite harboring the smallest genomes among plant RNA viruses, carmoviruses have emerged as an ideal model system for studying essential steps of the viral cycle including intracellular and intercellular trafficking. Two small movement proteins, formerly known as double gene block proteins (DGBp1 and DGBp2), have been involved in the movement throughout the plant of some members of carmovirus genera. DGBp1 RNA-binding capability was indispensable for cell-to-cell movement indicating that viral genomes must interact with DGBp1 to be transported. Further investigation on Melon necrotic spot virus (MNSV) DGBp1 subcellular localization and dynamics also supported this idea as this protein showed an actin-dependent movement along microfilaments and accumulated at the cellular periphery. Regarding DGBp2, subcellular localization studies showed that MNSV and Pelargonium flower break virus DGBp2s were inserted into the endoplasmic reticulum (ER) membrane but only MNSV DGBp2 trafficked to plasmodesmata (PD) via the Golgi apparatus through a COPII-dependent pathway. DGBp2 function is still unknown but its localization at PD was a requisite for an efficient cell-to-cell movement. It is also known that MNSV infection can induce a dramatic reorganization of mitochondria resulting in anomalous organelles containing viral RNAs. These putative viral factories were frequently found associated with the ER near the PD leading to the possibility that MNSV movement and replication could be spatially linked. Here, we update the current knowledge of the plant endomembrane system involvement in carmovirus intra- and intercellular movement and the tentative model proposed for MNSV transport within plant cells.
Keywords: Golgi; carmovirus; endoplasmic reticulum; intracellular movement; mitochondria; movement proteins.
Figures
References
-
- Blake J. A., Lee K. W., Morris T. J., Elthon T. E. (2007). Effects of Turnip crinkle virus infection on the structure and function of mitochondria and expression of stress proteins in turnips. Physiol. Plant. 129 698–706. 10.1111/j.1399-3054.2006.00852.x - DOI
-
- Blanco-Perez M., Perez-Canamas M., Ruiz L., Hernandez C. (2016). Efficient translation of Pelargonium line pattern virus RNAs relies on a TED-like 3′-translational enhancer that communicates with the corresponding 5′-region through a long-distance RNA-RNA interaction. PLOS ONE 11:e0152593. 10.1371/journal.pone.0152593 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources