

Substrates of the chloroplast small heat shock proteins 22E/F point to thermolability as a regulative switch for heat acclimation in Chlamydomonas reinhardtii
- PMID: 29094278
- PMCID: PMC5700999
- DOI: 10.1007/s11103-017-0672-y
Substrates of the chloroplast small heat shock proteins 22E/F point to thermolability as a regulative switch for heat acclimation in Chlamydomonas reinhardtii
Abstract
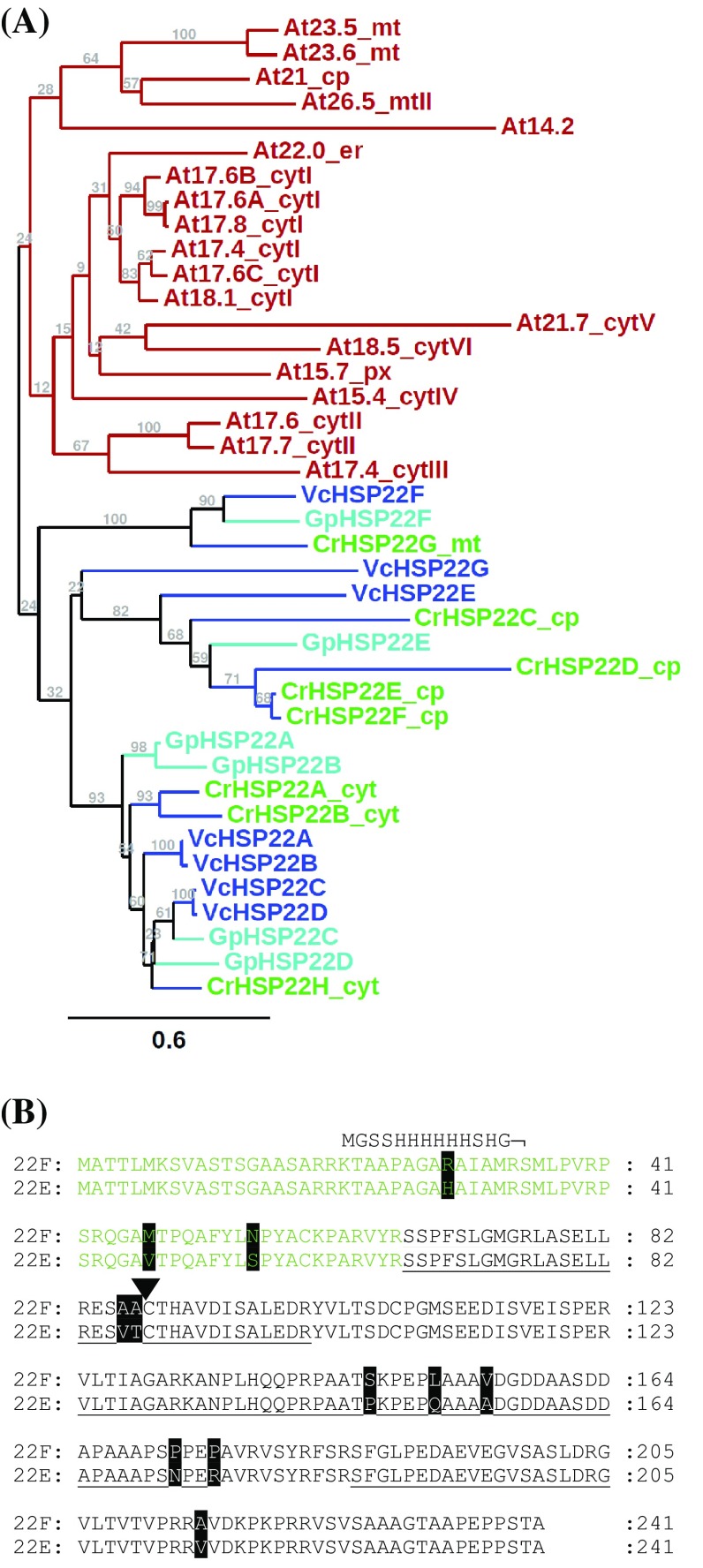
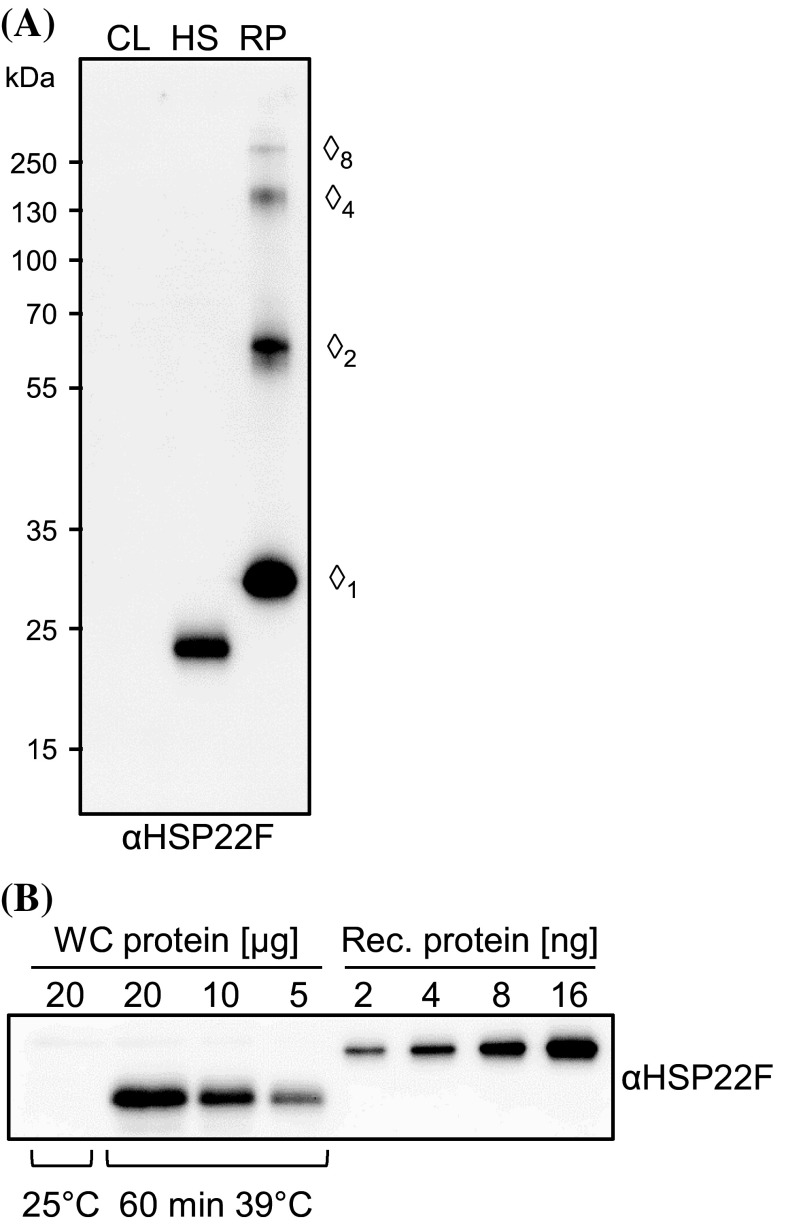
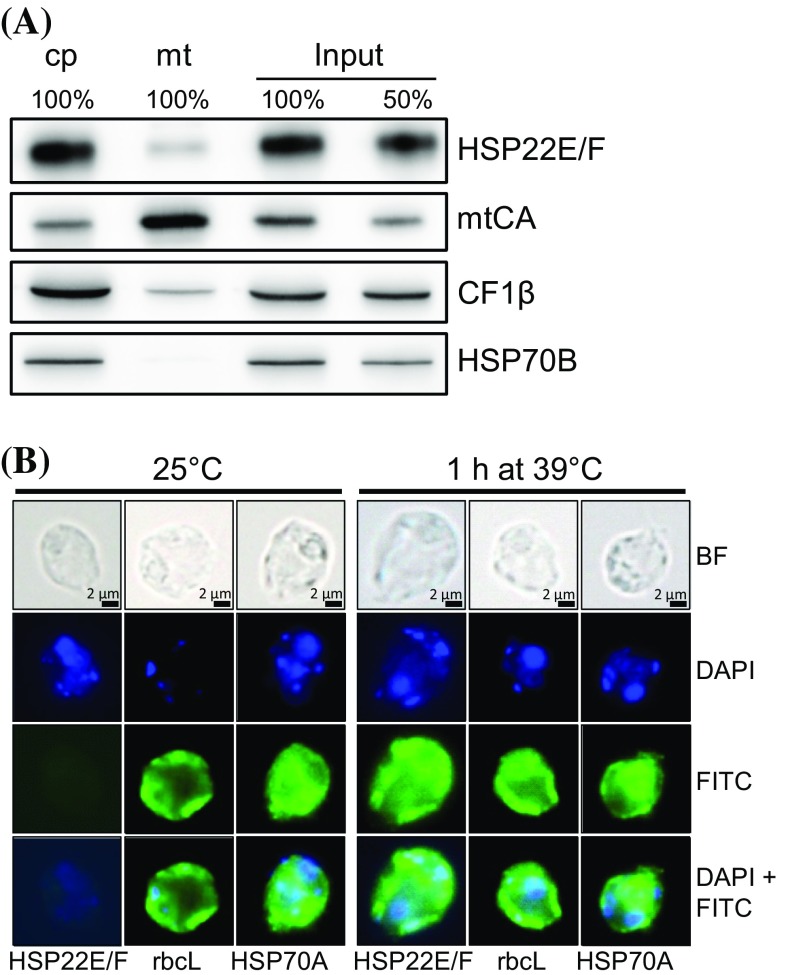
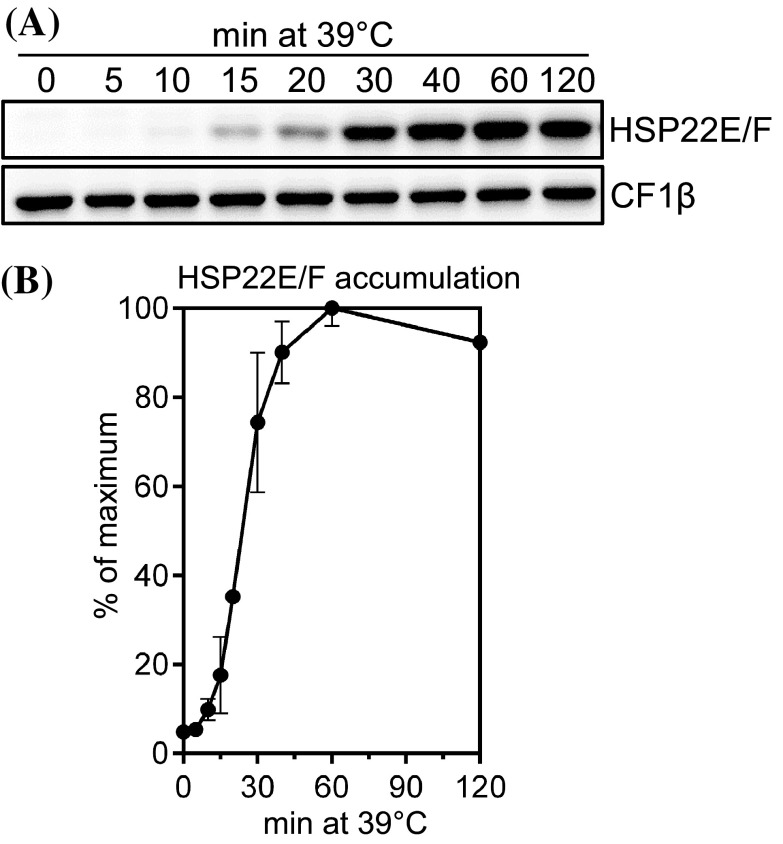
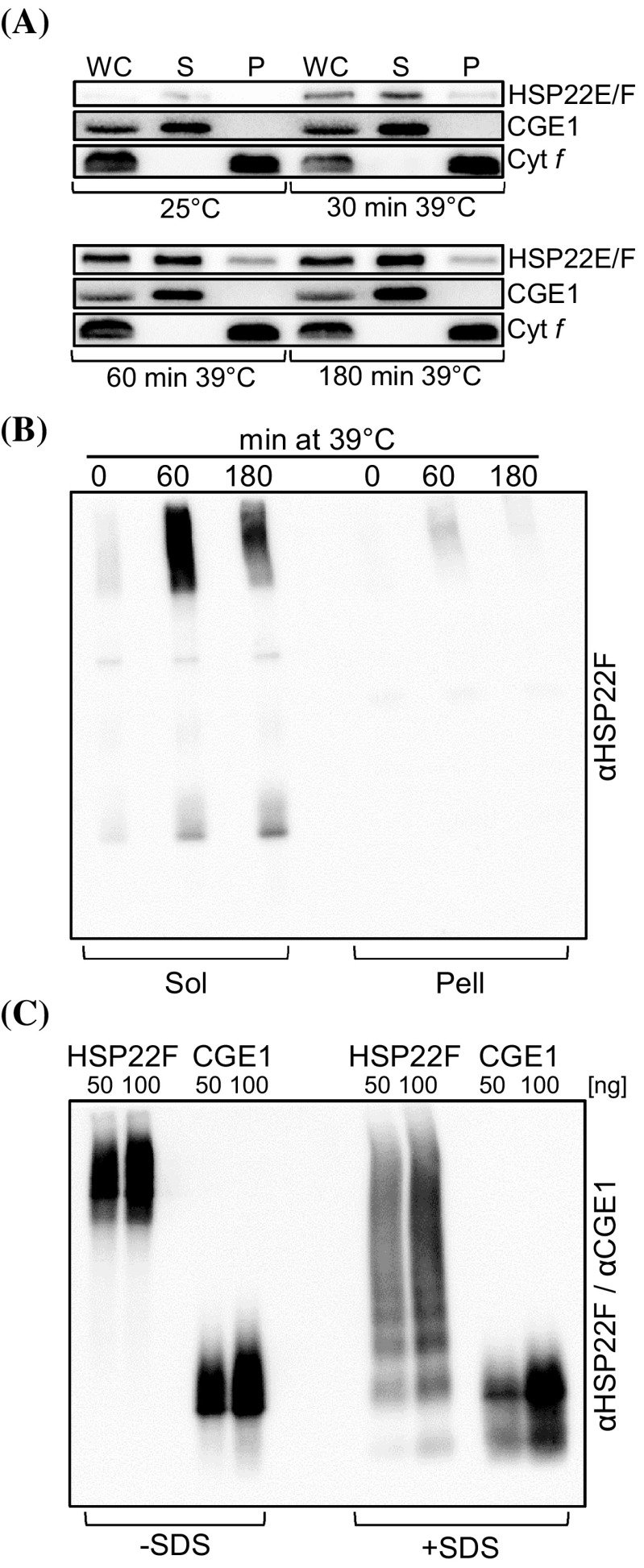
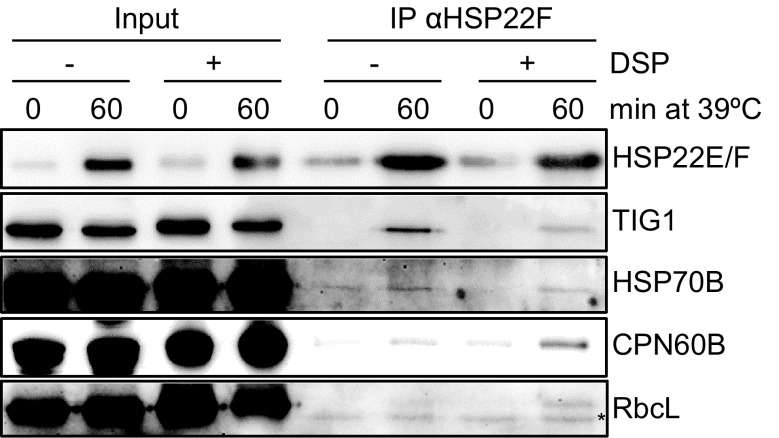
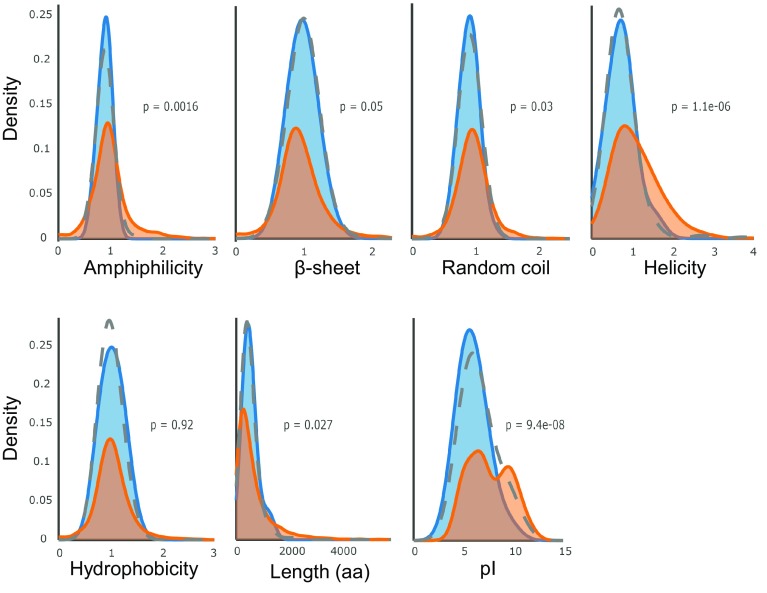
We have identified 39 proteins that interact directly or indirectly with high confidence with chloroplast HSP22E/F under heat stress thus revealing chloroplast processes affected by heat. Under conditions promoting protein unfolding, small heat shock proteins (sHsps) prevent the irreversible aggregation of unfolding proteins by integrating into forming aggregates. Aggregates containing sHsps facilitate the access of Hsp70 and ClpB/Hsp104 chaperones, which in ATP-dependent reactions disentangle individual proteins from the aggregates and assist in their refolding to the native state. Chlamydomonas reinhardtii encodes eight different sHsps (HSP22A to H). The goal of this work was to identify chloroplast-targeted sHsps in Chlamydomonas and to obtain a comprehensive list of the substrates with which they interact during heat stress in order to understand which chloroplast processes are disturbed under heat stress. We show that HSP22E and HSP22F are major chloroplast-targeted sHsps that have emerged from a recent gene duplication event resulting from the ongoing diversification of sHsps in the Volvocales. HSP22E/F strongly accumulate during heat stress and form high molecular mass complexes. Using differential immunoprecipitation, mass spectrometry and a stringent filtering algorithm we identified 39 proteins that with high-confidence interact directly or indirectly with HSP22E/F under heat stress. We propose that the apparent thermolability of several of these proteins might be a desired trait as part of a mechanism enabling Chlamydomonas chloroplasts to rapidly react to thermal stress.
Keywords: Chloroplast; Mass spectrometry; Molecular chaperones; Protein homeostasis; Protein–protein interactions.
Figures
References
-
- Basha E, Lee GJ, Breci LA, Hausrath AC, Buan NR, Giese KC, Vierling E. The identity of proteins associated with a small heat shock protein during heat stress in vivo indicates that these chaperones protect a wide range of cellular functions. J Biol Chem. 2004;279:7566–7575. doi: 10.1074/jbc.M310684200. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
