

Mitostasis in Neurons: Maintaining Mitochondria in an Extended Cellular Architecture
- PMID: 29096078
- PMCID: PMC5687842
- DOI: 10.1016/j.neuron.2017.09.055
Mitostasis in Neurons: Maintaining Mitochondria in an Extended Cellular Architecture
Abstract
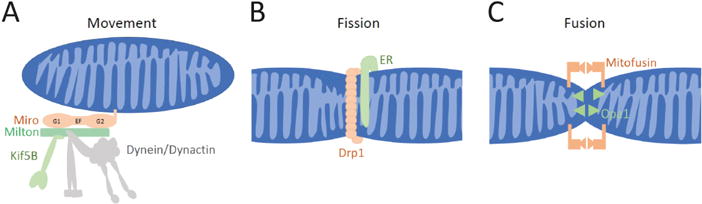
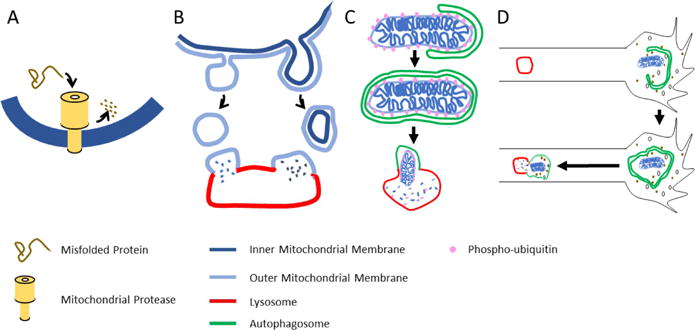
Neurons have more extended and complex shapes than other cells and consequently face a greater challenge in distributing and maintaining mitochondria throughout their arbors. Neurons can last a lifetime, but proteins turn over rapidly. Mitochondria, therefore, need constant rejuvenation no matter how far they are from the soma. Axonal transport of mitochondria and mitochondrial fission and fusion contribute to this rejuvenation, but local protein synthesis is also likely. Maintenance of a healthy mitochondrial population also requires the clearance of damaged proteins and organelles. This involves degradation of individual proteins, sequestration in mitochondria-derived vesicles, organelle degradation by mitophagy and macroautophagy, and in some cases transfer to glial cells. Both long-range transport and local processing are thus at work in achieving neuronal mitostasis-the maintenance of an appropriately distributed pool of healthy mitochondria for the duration of a neuron's life. Accordingly, defects in the processes that support mitostasis are significant contributors to neurodegenerative disorders.
Keywords: axonal transport; homeostasis; mitochondria; mitophagy; neurons.
Copyright © 2017 Elsevier Inc. All rights reserved.
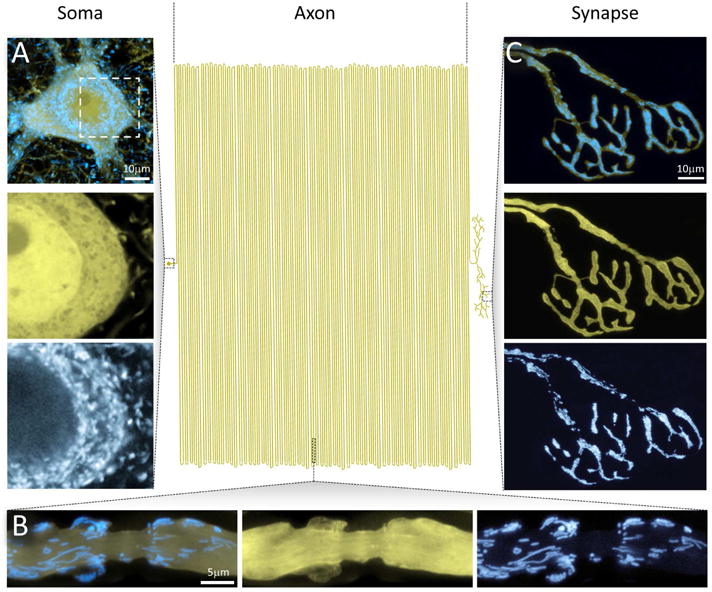
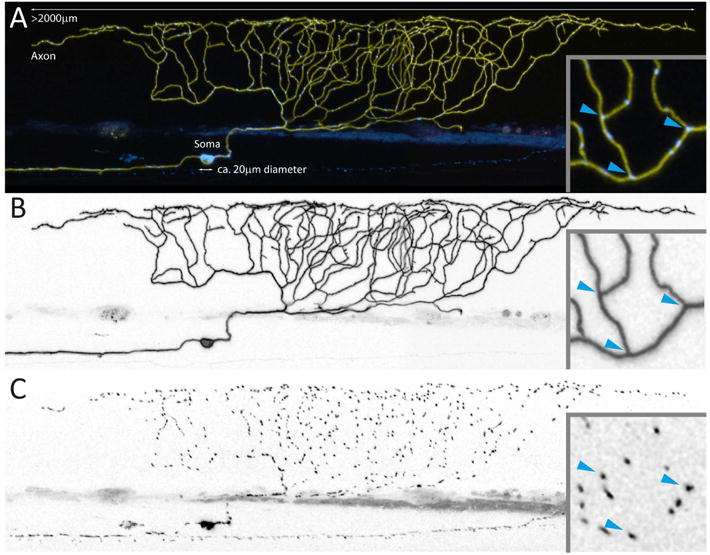
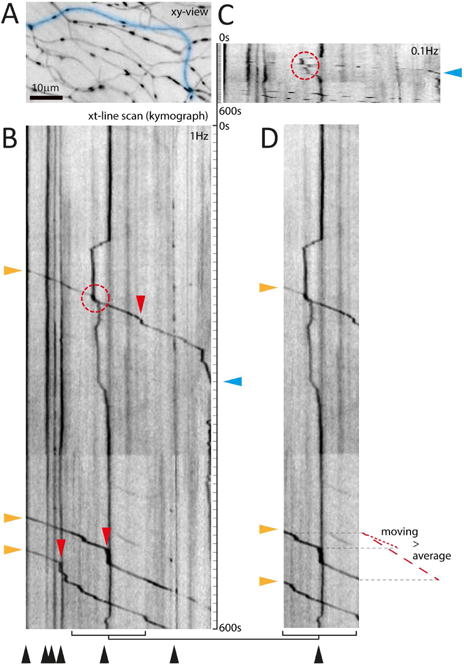
Figures
References
-
- Ahmed AU, Fisher PR. Import of nuclear-encoded mitochondrial proteins: a cotranslational perspective. Int Rev Cell Mol Biol. 2009;273:49–68. - PubMed
-
- Alexander C, Votruba M, Pesch UE, Thiselton DL, Mayer S, Moore A, Rodriguez M, Kellner U, Leo-Kottler B, Auburger G, et al. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat Genet. 2000;26:211–215. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
