

Spindle asymmetry drives non-Mendelian chromosome segregation
- PMID: 29097549
- PMCID: PMC5906099
- DOI: 10.1126/science.aan0092
Spindle asymmetry drives non-Mendelian chromosome segregation
Abstract
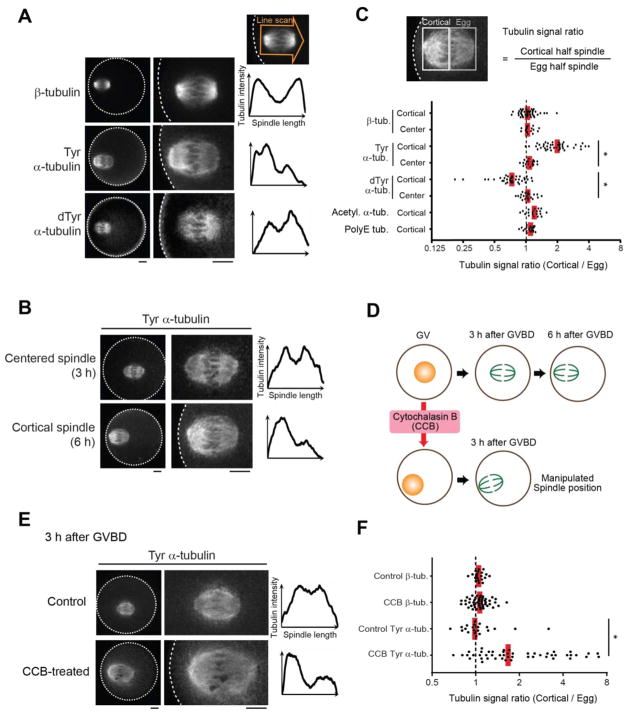
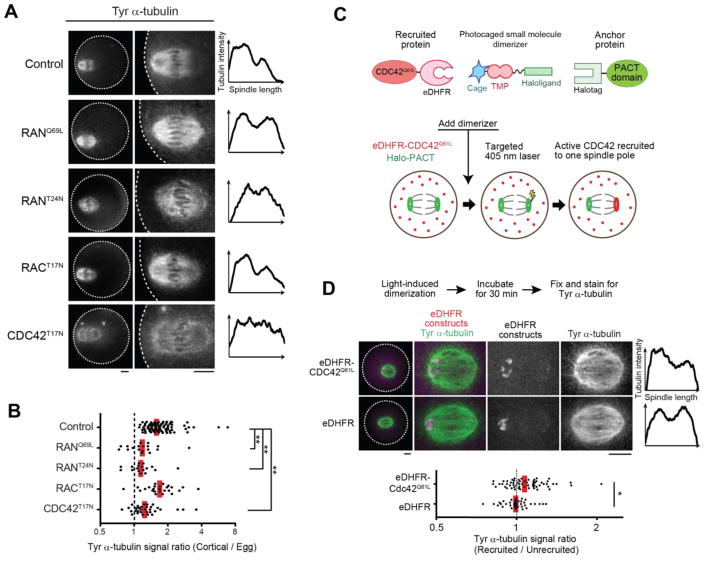
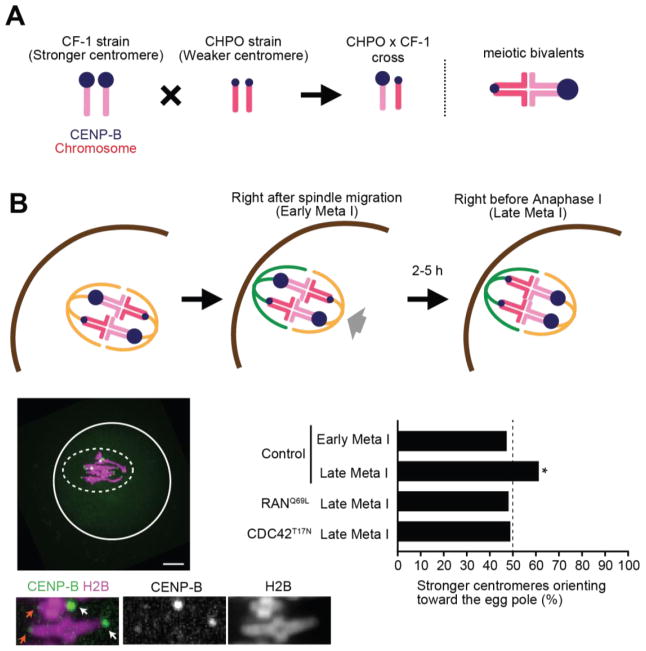
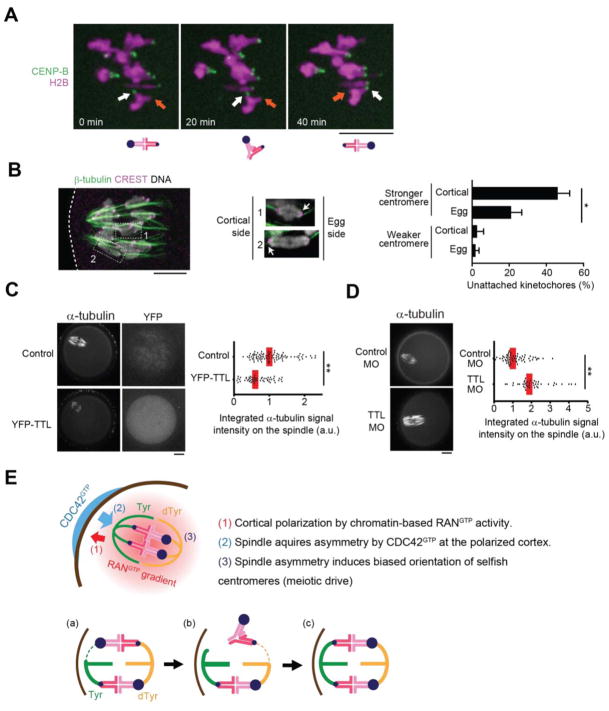
Genetic elements compete for transmission through meiosis, when haploid gametes are created from a diploid parent. Selfish elements can enhance their transmission through a process known as meiotic drive. In female meiosis, selfish elements drive by preferentially attaching to the egg side of the spindle. This implies some asymmetry between the two sides of the spindle, but the molecular mechanisms underlying spindle asymmetry are unknown. Here we found that CDC42 signaling from the cell cortex regulated microtubule tyrosination to induce spindle asymmetry and that non-Mendelian segregation depended on this asymmetry. Cortical CDC42 depends on polarization directed by chromosomes, which are positioned near the cortex to allow the asymmetric cell division. Thus, selfish meiotic drivers exploit the asymmetry inherent in female meiosis to bias their transmission.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
Comment in
-
Competing chromosomes explain junk DNA.Science. 2017 Nov 3;358(6363):594-595. doi: 10.1126/science.aaq0200. Science. 2017. PMID: 29097537 No abstract available.
-
Chromosome biology: Spindle asymmetry drives selfish segregation.Nat Rev Mol Cell Biol. 2018 Jan;19(1):2-3. doi: 10.1038/nrm.2017.121. Epub 2017 Nov 22. Nat Rev Mol Cell Biol. 2018. PMID: 29165428 No abstract available.
-
Asymmetric Tyrosination of Spindle Microtubules Facilitates Selfish Inheritance.Trends Cell Biol. 2018 Jun;28(6):417-419. doi: 10.1016/j.tcb.2018.03.005. Epub 2018 Apr 10. Trends Cell Biol. 2018. PMID: 29650359
References
-
- Rice WR. Nothing in Genetics Makes Sense Except in Light of Genomic Conflict. Annu Rev Ecol Evol Syst. 2013;44:217–237.
-
- Lindholm AK, et al. The Ecology and Evolutionary Dynamics of Meiotic Drive. Trends Ecol Evol. 2016;31:315–326. - PubMed
-
- Pardo-Manuel de Villena F, Sapienza C. Nonrandom segregation during meiosis: the unfairness of females. Mamm Genome. 2001;12:331–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
