

Mammalian γ2 AMPK regulates intrinsic heart rate
- PMID: 29097735
- PMCID: PMC5668267
- DOI: 10.1038/s41467-017-01342-5
Mammalian γ2 AMPK regulates intrinsic heart rate
Abstract
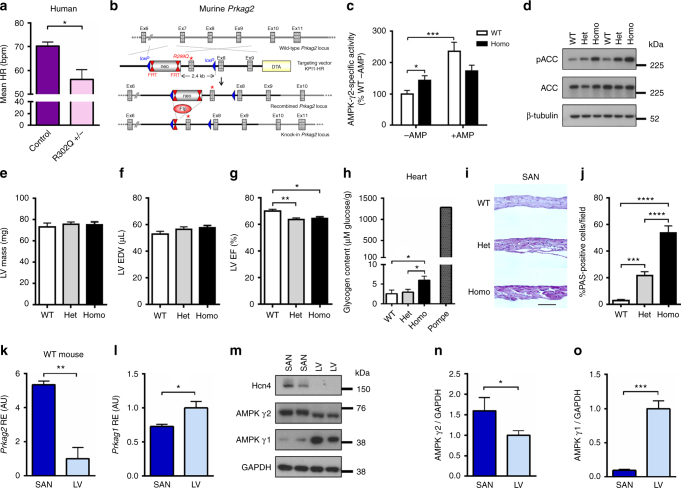
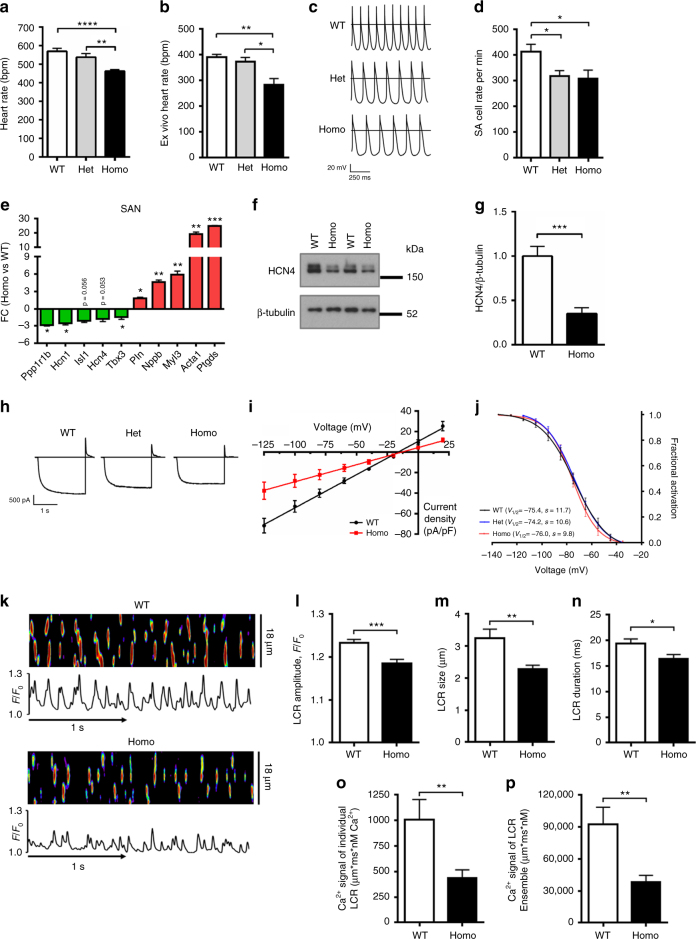
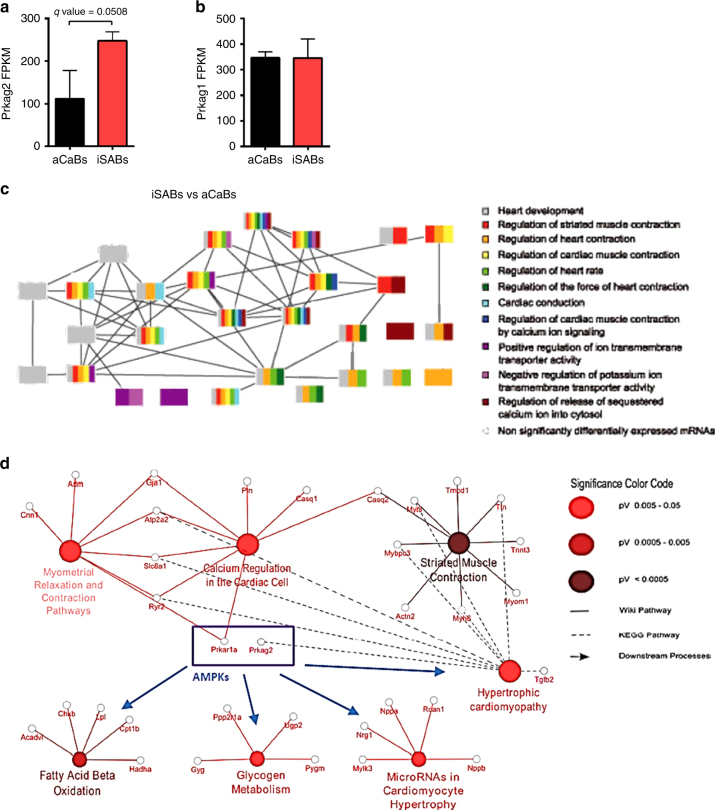
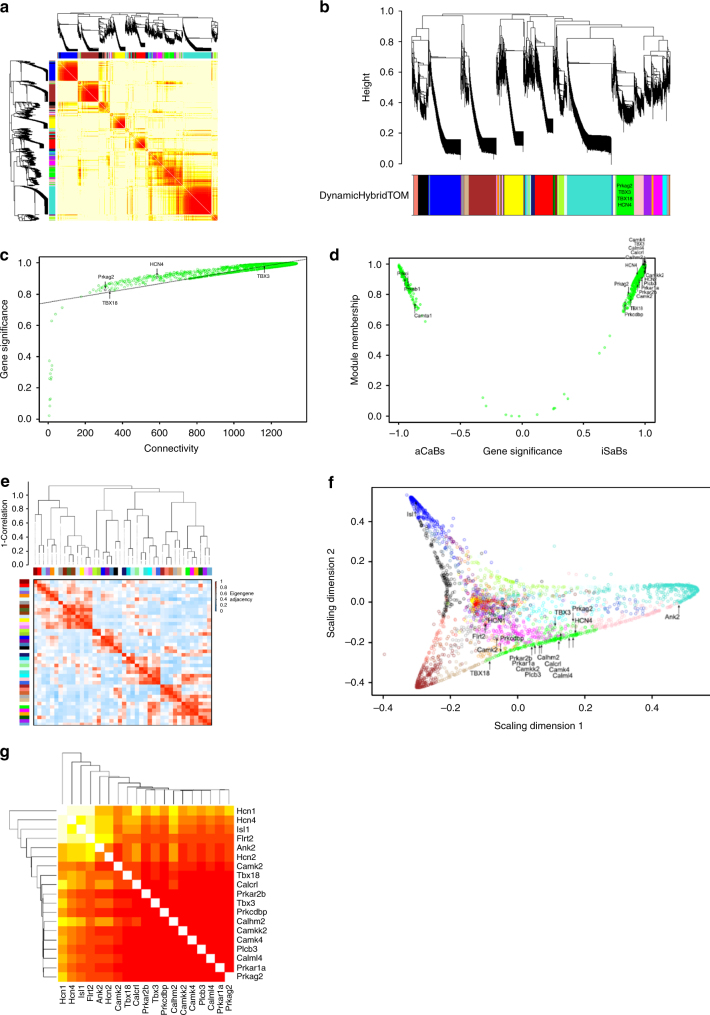
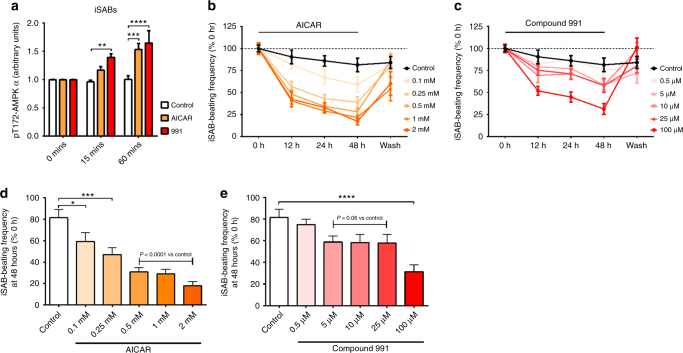
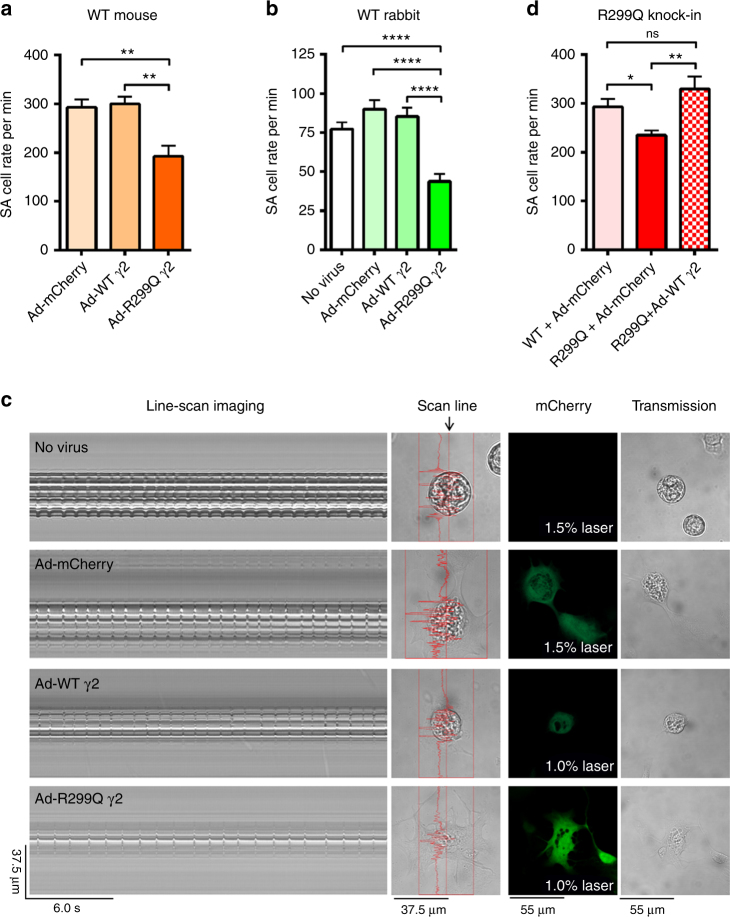
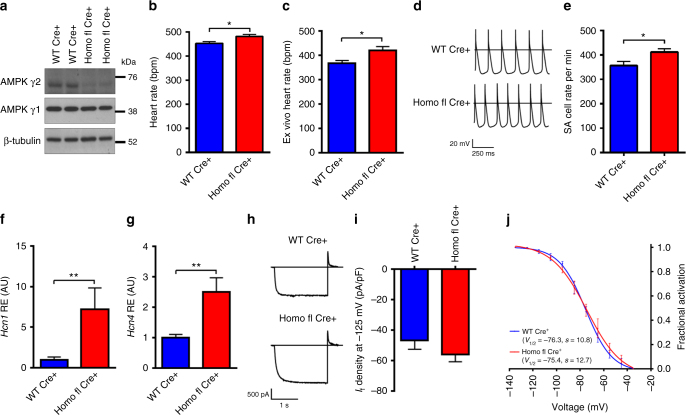
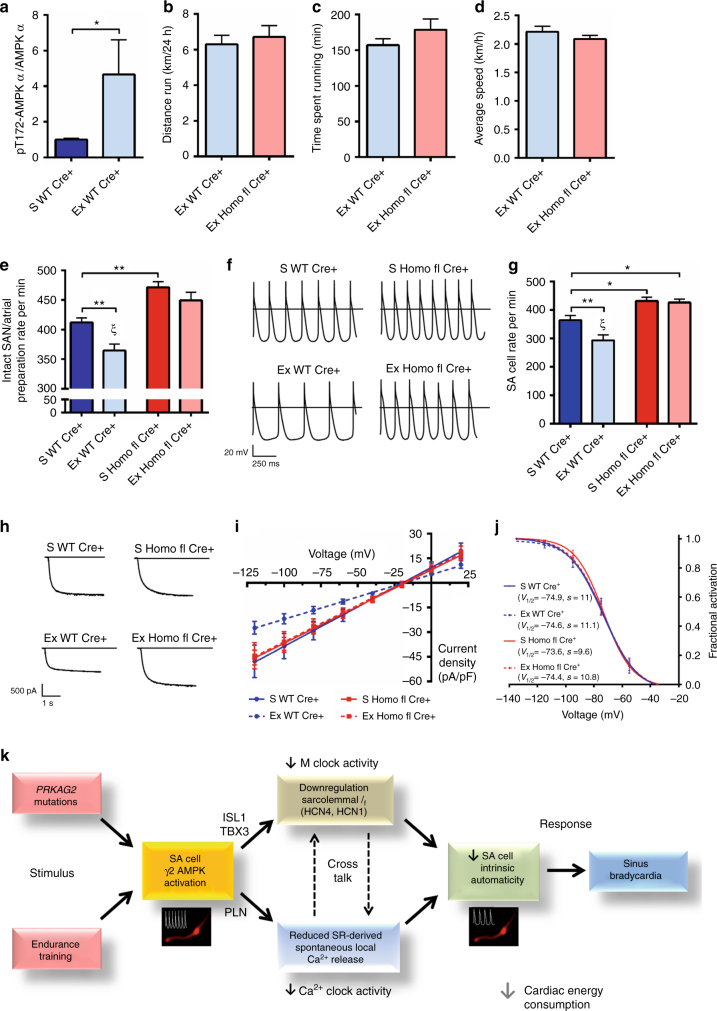
AMPK is a conserved serine/threonine kinase whose activity maintains cellular energy homeostasis. Eukaryotic AMPK exists as αβγ complexes, whose regulatory γ subunit confers energy sensor function by binding adenine nucleotides. Humans bearing activating mutations in the γ2 subunit exhibit a phenotype including unexplained slowing of heart rate (bradycardia). Here, we show that γ2 AMPK activation downregulates fundamental sinoatrial cell pacemaker mechanisms to lower heart rate, including sarcolemmal hyperpolarization-activated current (I f) and ryanodine receptor-derived diastolic local subsarcolemmal Ca2+ release. In contrast, loss of γ2 AMPK induces a reciprocal phenotype of increased heart rate, and prevents the adaptive intrinsic bradycardia of endurance training. Our results reveal that in mammals, for which heart rate is a key determinant of cardiac energy demand, AMPK functions in an organ-specific manner to maintain cardiac energy homeostasis and determines cardiac physiological adaptation to exercise by modulating intrinsic sinoatrial cell behavior.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- RG/07/012/24110/BHF_/British Heart Foundation/United Kingdom
- RG/12/5/29576/BHF_/British Heart Foundation/United Kingdom
- RG/12/16/29939/BHF_/British Heart Foundation/United Kingdom
- PG/14/41/30877/BHF_/British Heart Foundation/United Kingdom
- FS/11/50/29038/BHF_/British Heart Foundation/United Kingdom
- RG/07/003/23133/BHF_/British Heart Foundation/United Kingdom
- 086632/Z/08/Z/WT_/Wellcome Trust/United Kingdom
- R01 HL088427/HL/NHLBI NIH HHS/United States
- G0600829/MRC_/Medical Research Council/United Kingdom
- MC_UU_12010/6/MRC_/Medical Research Council/United Kingdom
- FS/15/8/31155/BHF_/British Heart Foundation/United Kingdom
- MC_U120027537/MRC_/Medical Research Council/United Kingdom
- MC_UU_00008/6/MRC_/Medical Research Council/United Kingdom
- G0200482/MRC_/Medical Research Council/United Kingdom
- RG/17/10/32859/BHF_/British Heart Foundation/United Kingdom
- FS/12/40/29712/BHF_/British Heart Foundation/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
