

Activation-induced cytidine deaminase prevents pro-B cell acute lymphoblastic leukemia by functioning as a negative regulator in Rag1 deficient pro-B cells
- PMID: 29100269
- PMCID: PMC5652663
- DOI: 10.18632/oncotarget.20563
Activation-induced cytidine deaminase prevents pro-B cell acute lymphoblastic leukemia by functioning as a negative regulator in Rag1 deficient pro-B cells
Abstract
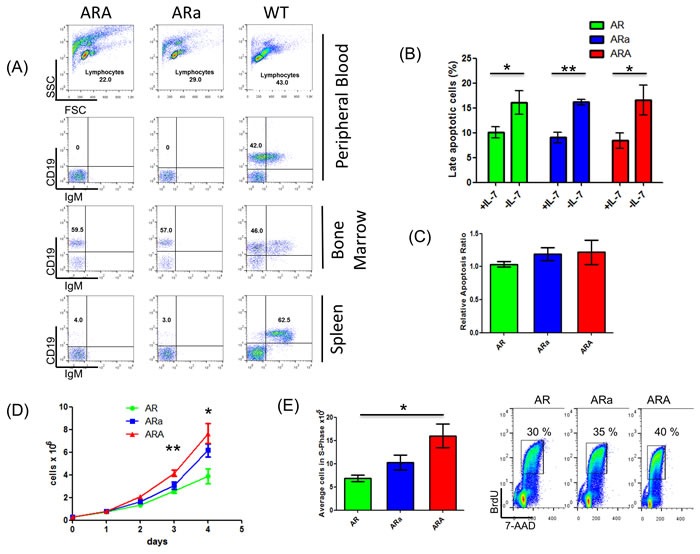
Activation-induced cytidine deaminase (AID) is essential for somatic hypermutation and class switch recombination in mature B-cells, while AID was also shown to play a role in developing pre-BCR/BCR-positive B-cells of the bone marrow. To further elucidate a potential function of Aid in the bone marrow prior to V(D)J-recombination, we utilized an in vivo model which exerts a B-cell developmental arrest at the pro-B cell stage with low frequencies of pro-B cell acute lymphoblastic leukemia (pro-B ALL) development. Therefore, p19Arf-/-Rag1-/- (AR) mice were crossed with Aid-deficient mice (ARA). Surprisingly, loss of Aid expression in pro-B cells accelerated pro-B ALL incidence from 30% (AR) to 98% (ARA). This effect was Aid dose dependent, since Aid+/- animals of the same background displayed a significantly lower incidence (83%). Furthermore, B-cell-specific Aid up-regulation was observed in Aid-competent pro-B ALLs. Additional whole exome/sanger sequencing of murine pro-B ALLs revealed an accumulation of recurrent somatic Jak3 (p.R653H, p.V670A) and Dnm2 (p.G397R) mutations, which highlights the importance of active IL7R signaling in the pro-B ALL blast cells. These findings were further supported by an enhanced proliferative potential of ARA pro-B cells compared to Aid-competent cells from the same genetic background. In summary, we show that both Aid and Rag1 act as a negative regulators in pro-B cells, preventing pro-B ALL.
Keywords: Rag1 deficiency; activation induced cytidine deaminase; acute lymphoblastic leukemia; pro-B cells.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no potential conflicts of interest.
Figures



Similar articles
-
Activation-induced cytidine deaminase accelerates clonal evolution in BCR-ABL1-driven B-cell lineage acute lymphoblastic leukemia.Cancer Res. 2010 Oct 1;70(19):7411-20. doi: 10.1158/0008-5472.CAN-10-1438. Epub 2010 Sep 28. Cancer Res. 2010. PMID: 20876806 Free PMC article.
-
Activation-induced cytidine deaminase acts as a mutator in BCR-ABL1-transformed acute lymphoblastic leukemia cells.J Exp Med. 2007 May 14;204(5):1157-66. doi: 10.1084/jem.20062662. Epub 2007 May 7. J Exp Med. 2007. PMID: 17485517 Free PMC article.
-
AID-induced T-lymphoma or B-leukemia/lymphoma in a mouse BMT model.Leukemia. 2010 May;24(5):1018-24. doi: 10.1038/leu.2010.40. Epub 2010 Apr 1. Leukemia. 2010. PMID: 20357822
-
Activation-induced cytidine deaminase: a dual role in class-switch recombination and somatic hypermutation.Eur J Immunol. 2003 Aug;33(8):2069-73. doi: 10.1002/eji.200324133. Eur J Immunol. 2003. PMID: 12884279 Review.
-
Epigenetic function of activation-induced cytidine deaminase and its link to lymphomagenesis.Front Immunol. 2014 Dec 18;5:642. doi: 10.3389/fimmu.2014.00642. eCollection 2014. Front Immunol. 2014. PMID: 25566255 Free PMC article. Review.
Cited by
-
Transcriptional Regulation of Genes by Ikaros Tumor Suppressor in Acute Lymphoblastic Leukemia.Int J Mol Sci. 2020 Feb 18;21(4):1377. doi: 10.3390/ijms21041377. Int J Mol Sci. 2020. PMID: 32085659 Free PMC article. Review.
-
Frequent expression of activation-induced cytidine deaminase in diffuse large B-cell lymphoma tissues from persons living with HIV.AIDS. 2020 Nov 15;34(14):2025-2035. doi: 10.1097/QAD.0000000000002653. AIDS. 2020. PMID: 32773475 Free PMC article.
References
-
- Zhang J, McCastlain K, Yoshihara H, Xu B, Chang Y, Churchman ML, Wu G, Li Y, Wei L, Iacobucci I, Liu Y, Qu C, Wen J, et al. Deregulation of DUX4 and ERG in acute lymphoblastic leukemia. Nat Genet. 2016;48:1481–9. http://doi.org/10.1038/ng.3691. - DOI - PMC - PubMed
-
- Hunger SP, Mullighan CG. Acute Lymphoblastic Leukemia in Children. N Engl J Med. 2015;373:1541–52. http://doi.org/10.1056/NEJMra1400972. - DOI - PubMed
-
- Pui CH, Yang JJ, Hunger SP, Pieters R, Schrappe M, Biondi A, Vora A, Baruchel A, Silverman LB, Schmiegelow K, Escherich G, Horibe K, Benoit YC, et al. Childhood Acute Lymphoblastic Leukemia: Progress Through Collaboration. J Clin Oncol. 2015;33:2938–48. http://doi.org/10.1200/JCO.2014.59.1636. - DOI - PMC - PubMed
-
- Mullighan CG, Goorha S, Radtke I, Miller CB, Coustan-Smith E, Dalton JD, Girtman K, Mathew S, Ma J, Pounds SB, Su X, Pui CH, Relling MV, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature. 2007;446:758–64. http://doi.org/10.1038/nature05690. - DOI - PubMed
-
- Shah S, Schrader KA, Waanders E, Timms AE, Vijai J, Miething C, Wechsler J, Yang J, Hayes J, Klein RJ, Zhang J, Wei L, Wu G, et al. A recurrent germline PAX5 mutation confers susceptibility to pre-B cell acute lymphoblastic leukemia. Nat Genet. 2013;45:1226–31. http://doi.org/10.1038/ng.2754. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
