

A tyrosine kinase-STAT5-miR21-PDCD4 regulatory axis in chronic and acute myeloid leukemia cells
- PMID: 29100302
- PMCID: PMC5652696
- DOI: 10.18632/oncotarget.19192
A tyrosine kinase-STAT5-miR21-PDCD4 regulatory axis in chronic and acute myeloid leukemia cells
Abstract
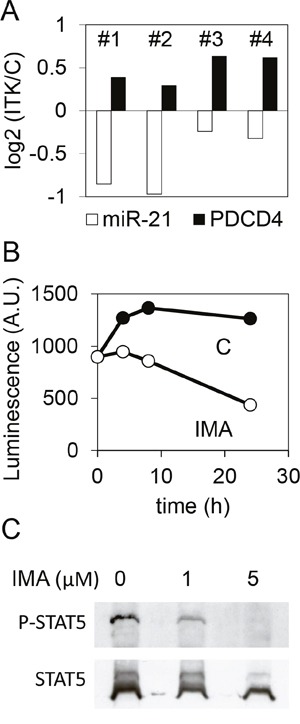
MicroRNAs (miRNAs) are regulators of several key patho-physiological processes, including cell cycle and apoptosis. Using microarray-based miRNA profiling in K562 cells, a model of chronic myeloid leukemia (CML), we found that the oncoprotein BCR-ABL1 regulates the expression of miR-21, an "onco-microRNA", found to be overexpressed in several cancers. This effect relies on the presence of two STAT binding sites on the promoter of miR-21, and on the phosphorylation status of STAT5, a transcription factor activated by the kinase activity of BCR-ABL1. Mir-21 regulates the expression of PDCD4 (programmed cell death protein 4), a tumor suppressor identified through a proteomics approach. The phosphoSTAT5 - miR-21 - PDCD4 pathway was active in CML primary CD34+ cells, but also in acute myeloid leukemia (AML) models like MV4.11 and MOLM13, where the constitutively active tyrosine kinase FLT3-ITD plays a similar role to BCR-ABL1 in the K562 cell line.
Keywords: CML; STAT5; leukemia; miR-21; microRNA.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare no conflicts of interest
Figures







References
-
- Jacquel A, Herrant M, Legros L, Belhacene N, Luciano F, Pages G, Hofman P, Auberger P. Imatinib induces mitochondria-dependent apoptosis of the Bcr-Abl-positive K562 cell line and its differentiation toward the erythroid lineage. FASEB J Off Publ Fed Am Soc Exp Biol. 2003;17:2160–2. doi: 10.1096/fj.03-0322. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
