

Cellular, ultrastructural and molecular analyses of epidermal cell development in the planarian Schmidtea mediterranea
- PMID: 29100657
- PMCID: PMC5750087
- DOI: 10.1016/j.ydbio.2017.08.030
Cellular, ultrastructural and molecular analyses of epidermal cell development in the planarian Schmidtea mediterranea
Abstract
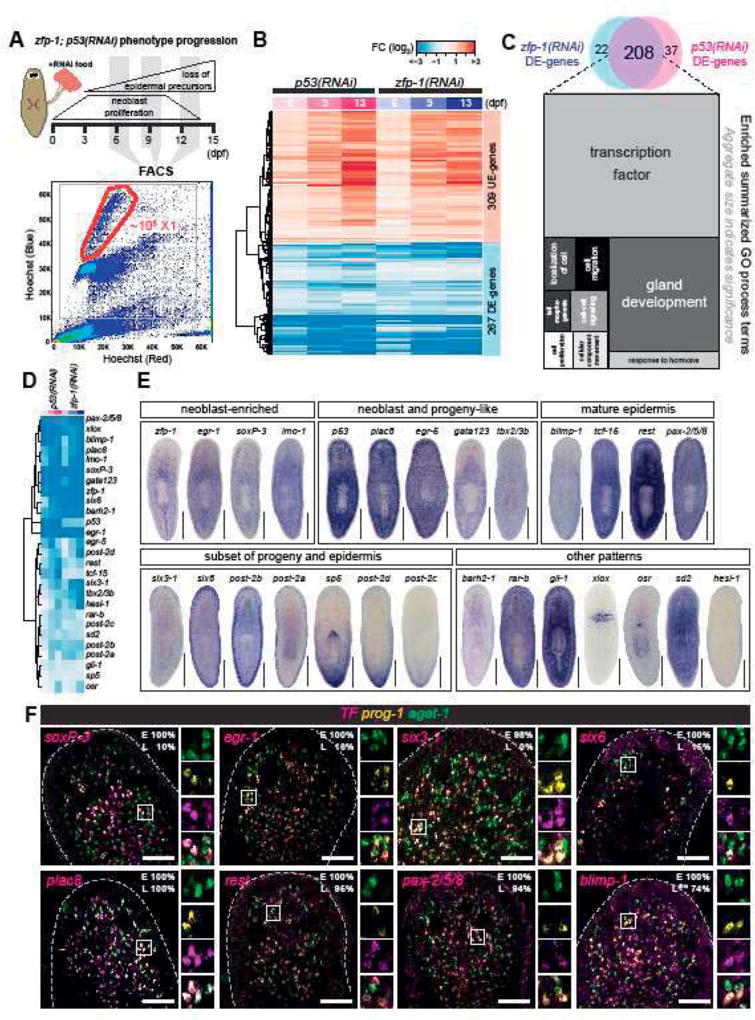
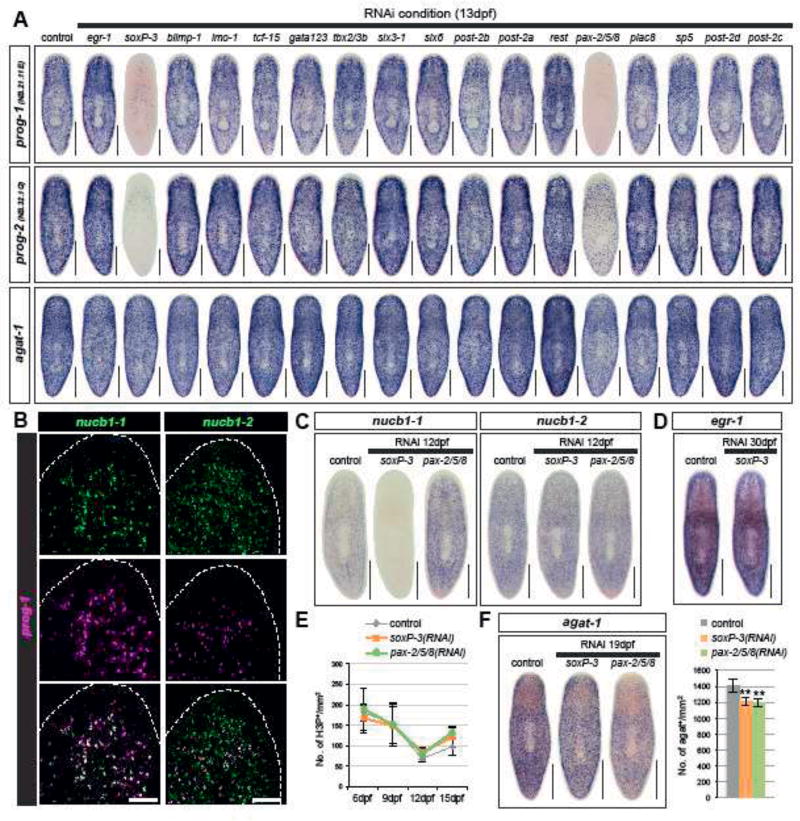
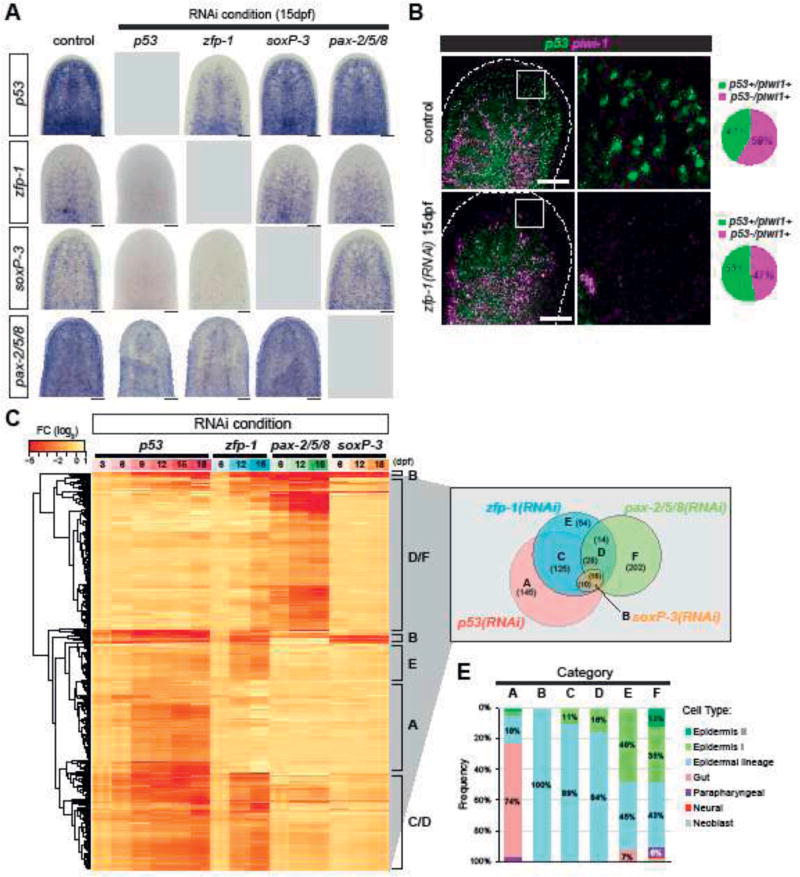
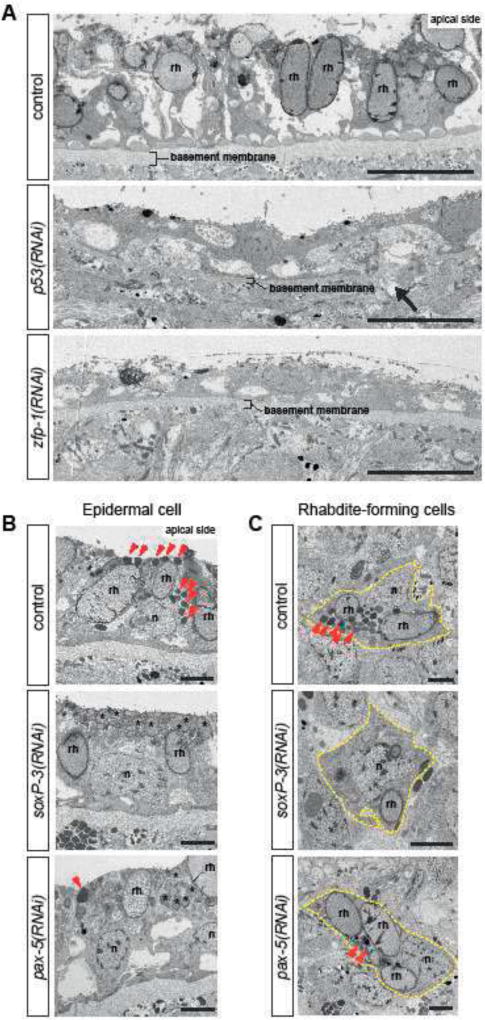
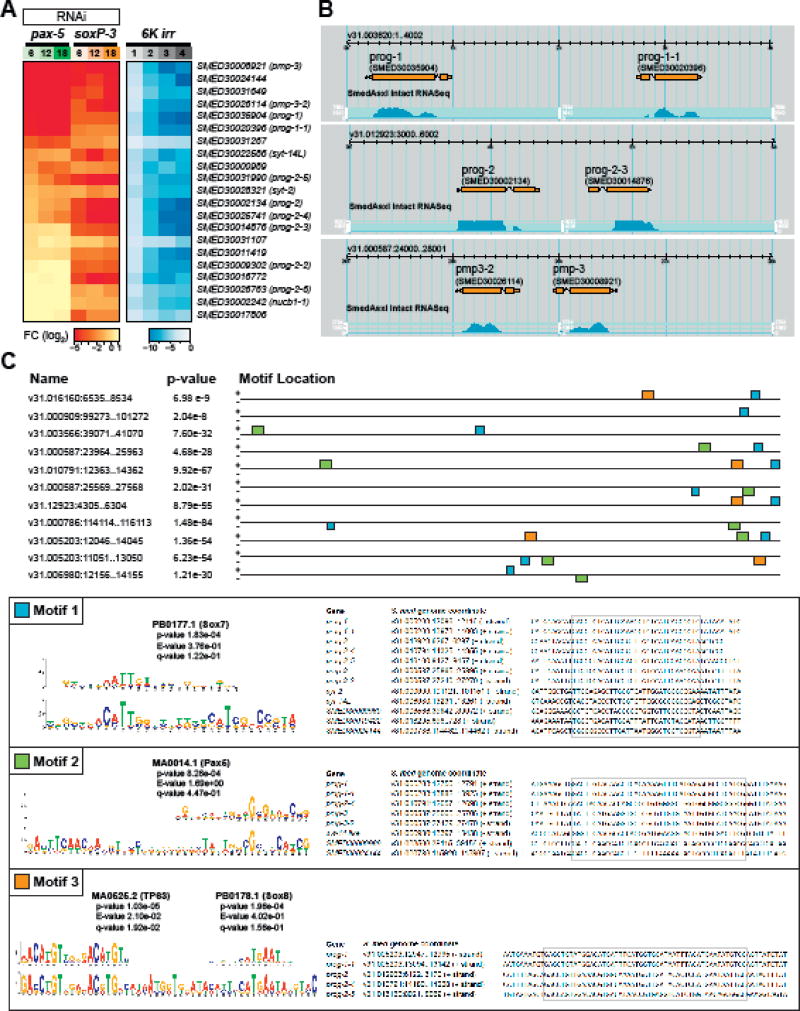
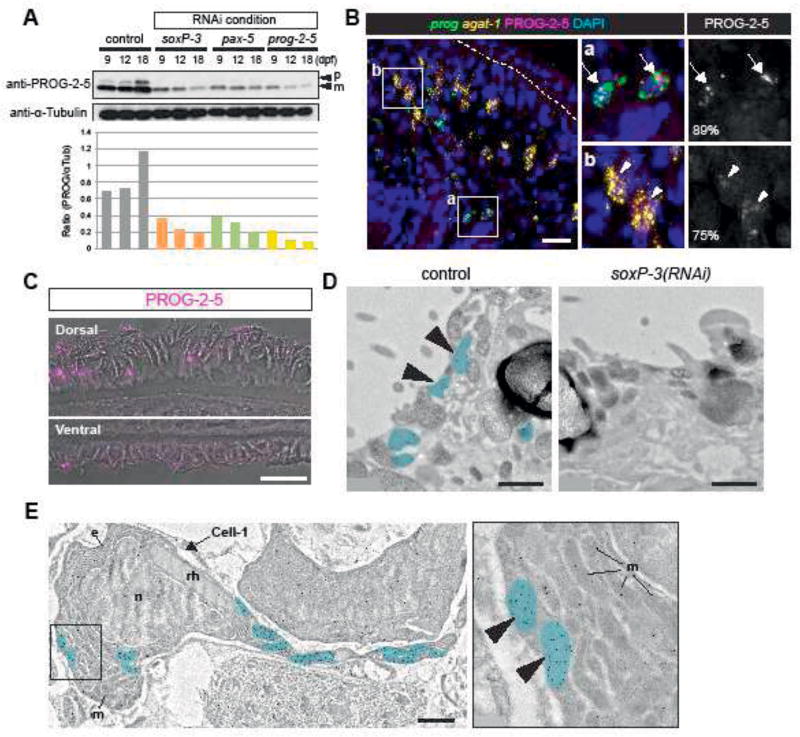
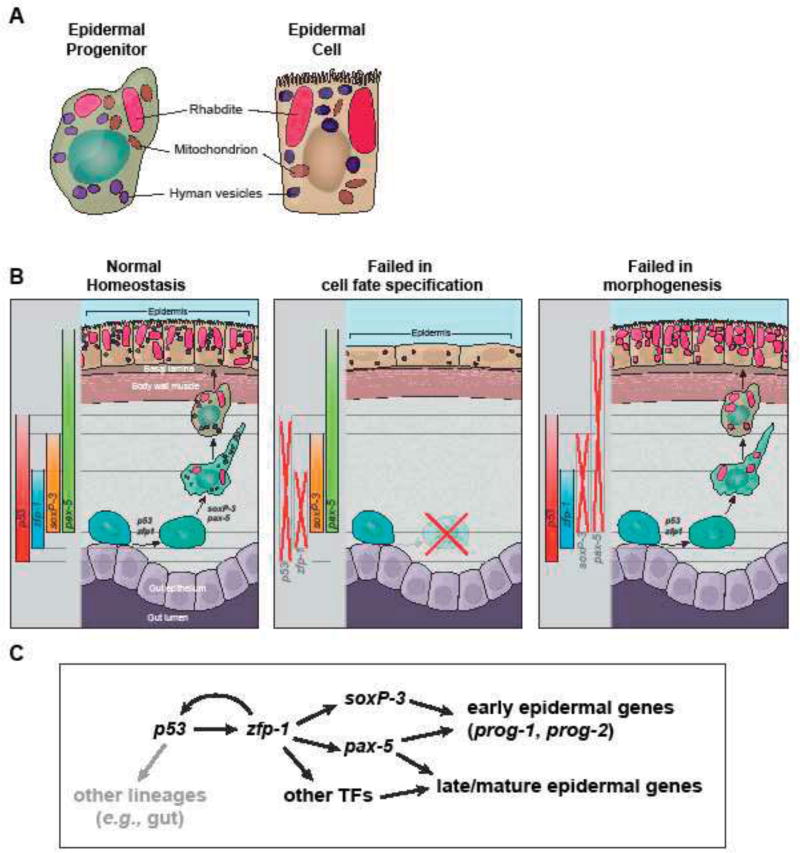
The epidermis is essential for animal survival, providing both a protective barrier and cellular sensor to external environments. The generally conserved embryonic origin of the epidermis, but the broad morphological and functional diversity of this organ across animals is puzzling. We define the transcriptional regulators underlying epidermal lineage differentiation in the planarian Schmidtea mediterranea, an invertebrate organism that, unlike fruitflies and nematodes, continuously replaces its epidermal cells. We find that Smed-p53, Sox and Pax transcription factors are essential regulators of epidermal homeostasis, and act cooperatively to regulate genes associated with early epidermal precursor cell differentiation, including a tandemly arrayed novel gene family (prog) of secreted proteins. Additionally, we report on the discovery of distinct and previously undescribed secreted organelles whose production is dependent on the transcriptional activity of soxP-3, and which we term Hyman vesicles.
Keywords: Epidermis; Stem cells; Transcription; p53; pax-2/5/8; soxP-3; zfp-1.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Benjamini Y, Hochberg Y. On the Adaptive Control of the False Discovery Rate in Multiple Testing with Independent Statistics. Journal of Educational and Behavioral Statistics. 2000;25(1):60.
-
- Cebrià F, Newmark PA. Planarian homologs of netrin and netrin receptor are required for proper regeneration of the central nervous system and the maintenance of nervous system architecture. Development. 2005;132(16):3691–3703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
