

Tinkering with Osmotically Controlled Transcription Allows Enhanced Production and Excretion of Ectoine and Hydroxyectoine from a Microbial Cell Factory
- PMID: 29101191
- PMCID: PMC5752866
- DOI: 10.1128/AEM.01772-17
Tinkering with Osmotically Controlled Transcription Allows Enhanced Production and Excretion of Ectoine and Hydroxyectoine from a Microbial Cell Factory
Abstract
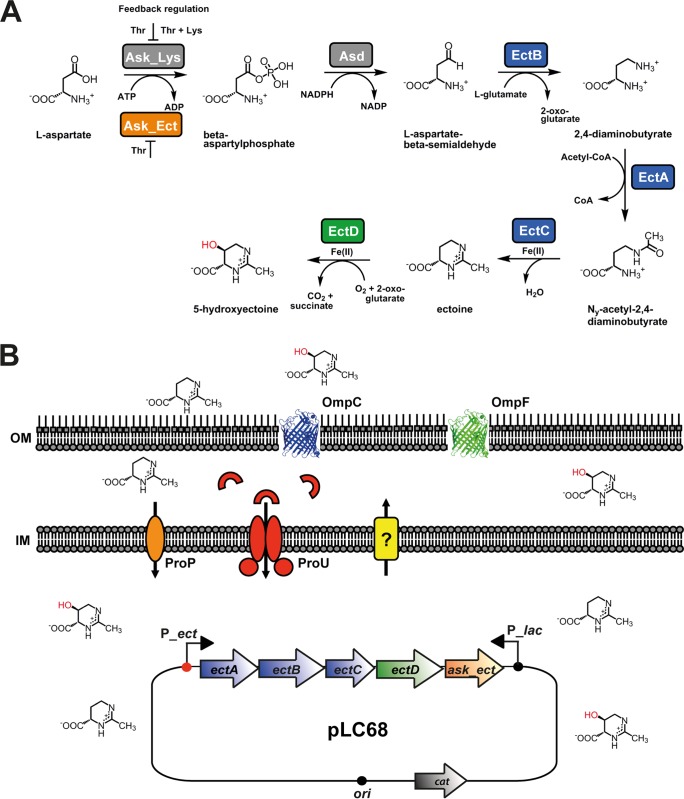
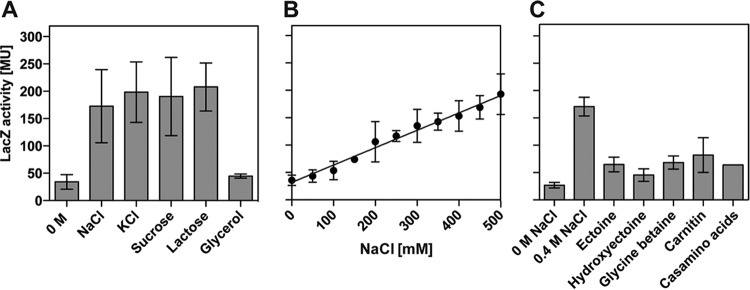
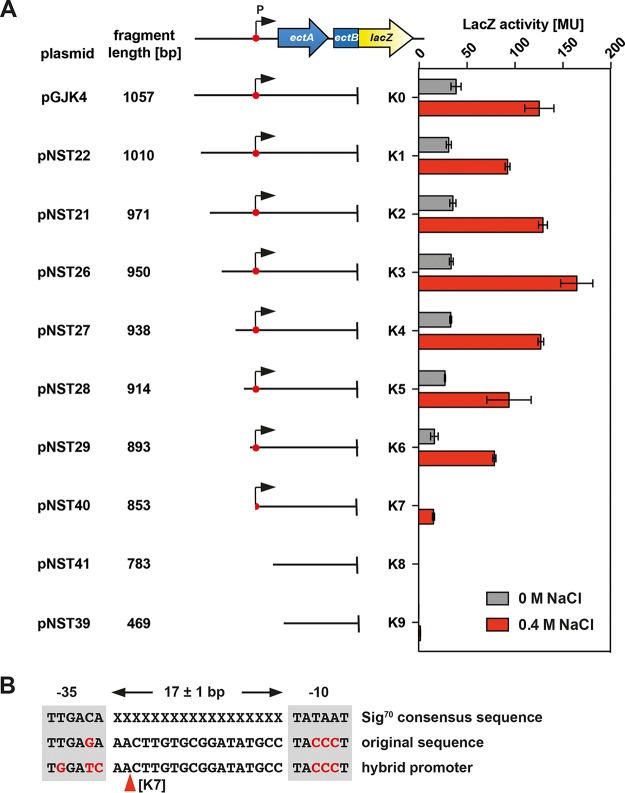
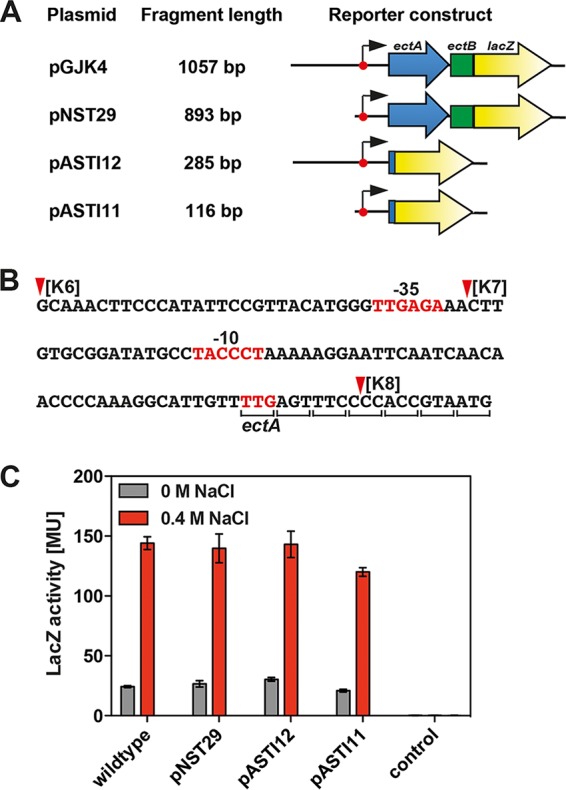
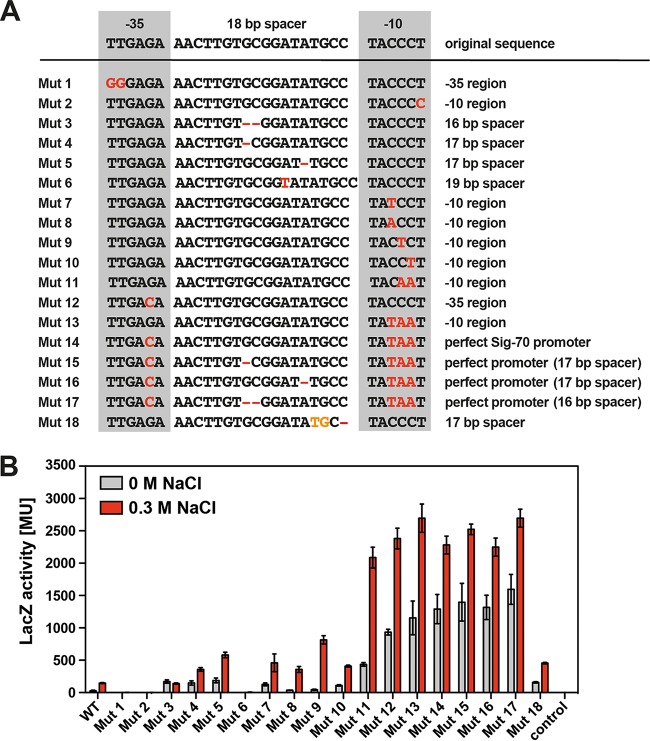
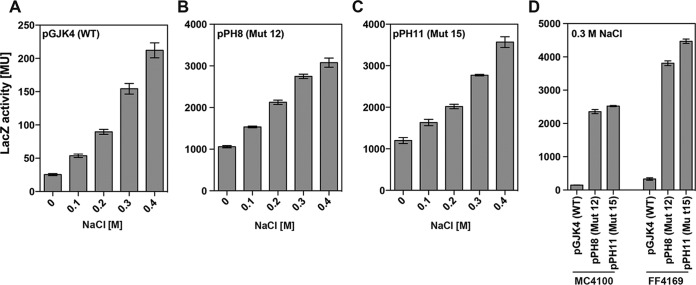
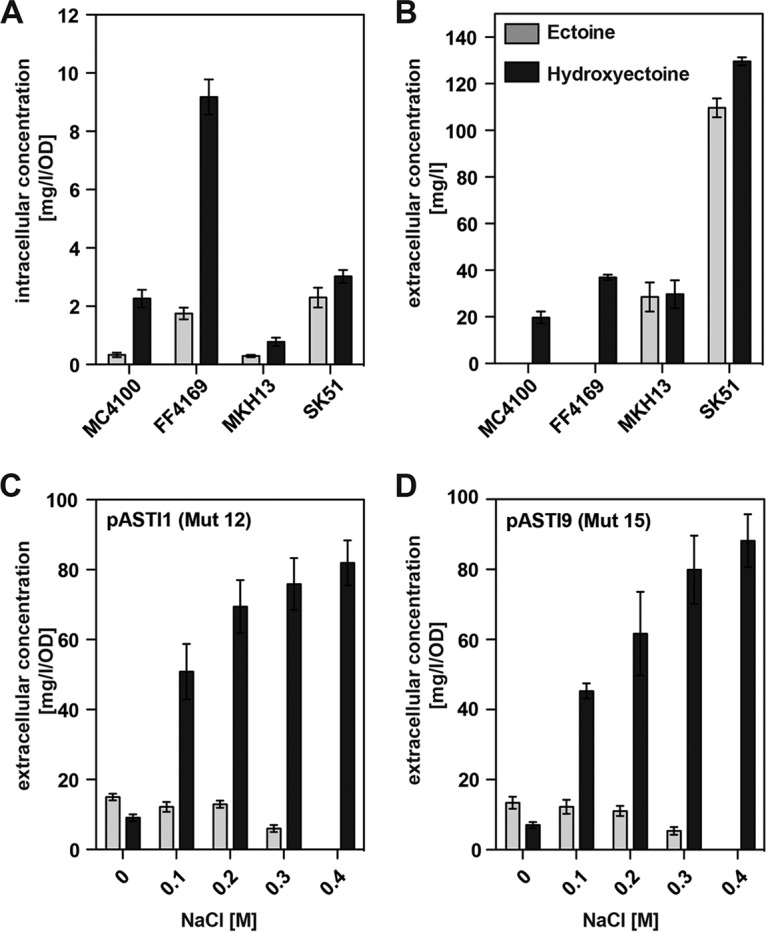
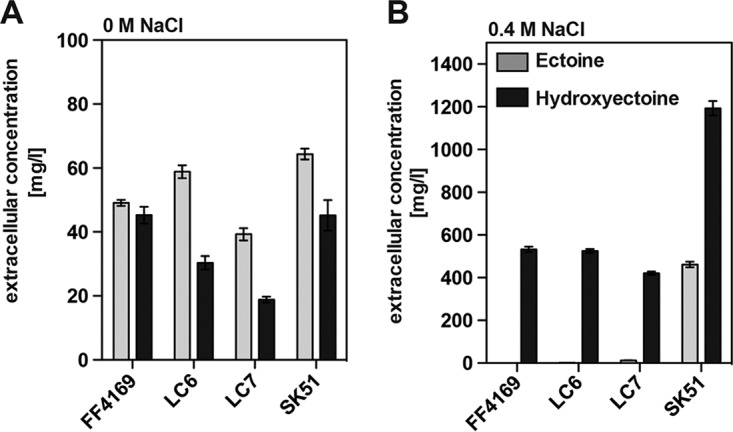
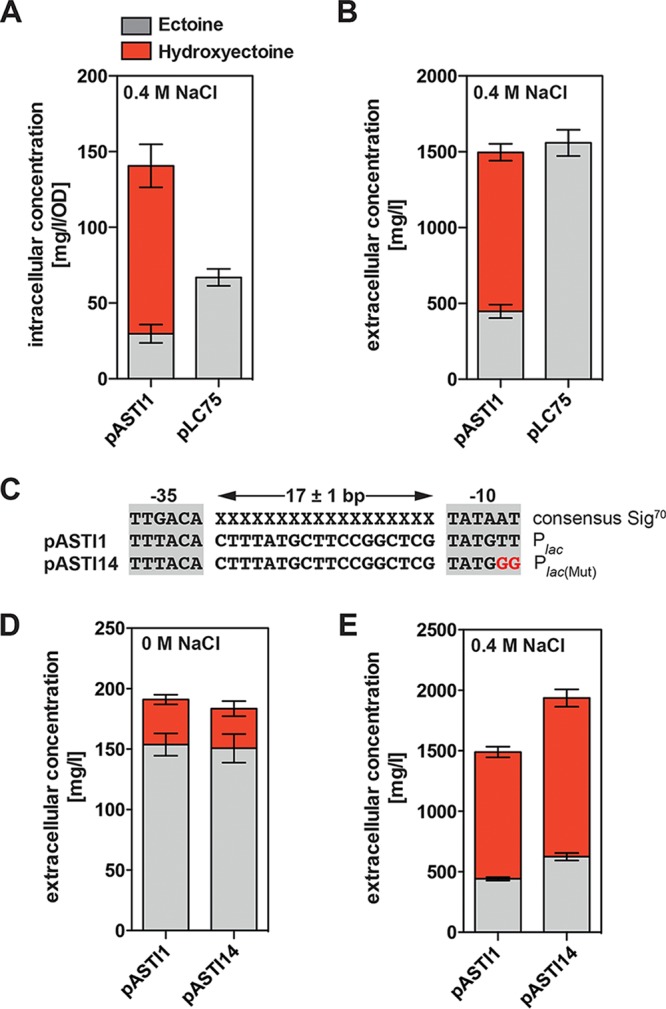
Ectoine and hydroxyectoine are widely synthesized by members of the Bacteria and a few members of the Archaea as potent osmostress protectants. We have studied the salient features of the osmostress-responsive promoter directing the transcription of the ectoine/hydroxyectoine biosynthetic gene cluster from the plant-root-associated bacterium Pseudomonas stutzeri by transferring it into Escherichia coli, an enterobacterium that does not produce ectoines naturally. Using ect-lacZ reporter fusions, we found that the heterologous ect promoter reacted with exquisite sensitivity in its transcriptional profile to graded increases in sustained high salinity, responded to a true osmotic signal, and required the buildup of an osmotically effective gradient across the cytoplasmic membrane for its induction. The involvement of the -10, -35, and spacer regions of the sigma-70-type ect promoter in setting promoter strength and response to osmotic stress was assessed through site-directed mutagenesis. Moderate changes in the ect promoter sequence that increase its resemblance to housekeeping sigma-70-type promoters of E. coli afforded substantially enhanced expression, both in the absence and in the presence of osmotic stress. Building on this set of ect promoter mutants, we engineered an E. coli chassis strain for the heterologous production of ectoines. This synthetic cell factory lacks the genes for the osmostress-responsive synthesis of trehalose and the compatible solute importers ProP and ProU, and it continuously excretes ectoines into the growth medium. By combining appropriate host strains and different plasmid variants, excretion of ectoine, hydroxyectoine, or a mixture of both compounds was achieved under mild osmotic stress conditions.IMPORTANCE Ectoines are compatible solutes, organic osmolytes that are used by microorganisms to fend off the negative consequences of high environmental osmolarity on cellular physiology. An understanding of the salient features of osmostress-responsive promoters directing the expression of the ectoine/hydroxyectoine biosynthetic gene clusters is lacking. We exploited the ect promoter from an ectoine/hydroxyectoine-producing soil bacterium for such a study by transferring it into a surrogate bacterial host. Despite the fact that E. coli does not synthesize ectoines naturally, the ect promoter retained its exquisitely sensitive osmotic control, indicating that osmoregulation of ect transcription is an inherent feature of the promoter and its flanking sequences. These sequences were narrowed to a 116-bp DNA fragment. Ectoines have interesting commercial applications. Building on data from a site-directed mutagenesis study of the ect promoter, we designed a synthetic cell factory that secretes ectoine, hydroxyectoine, or a mixture of both compounds into the growth medium.
Keywords: chemical chaperones; compatible solutes; excretion; osmoregulation; promoters.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Widderich N, Czech L, Elling FJ, Könneke M, Stöveken N, Pittelkow M, Riclea R, Dickschat JS, Heider J, Bremer E. 2016. Strangers in the archaeal world: osmostress-responsive biosynthesis of ectoine and hydroxyectoine by the marine thaumarchaeon Nitrosopumilus maritimus. Environ Microbiol 18:1227–1248. doi: 10.1111/1462-2920.13156. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
