

TFIIH generates a six-base-pair open complex during RNAP II transcription initiation and start-site scanning
- PMID: 29106413
- PMCID: PMC5741190
- DOI: 10.1038/nsmb.3500
TFIIH generates a six-base-pair open complex during RNAP II transcription initiation and start-site scanning
Abstract
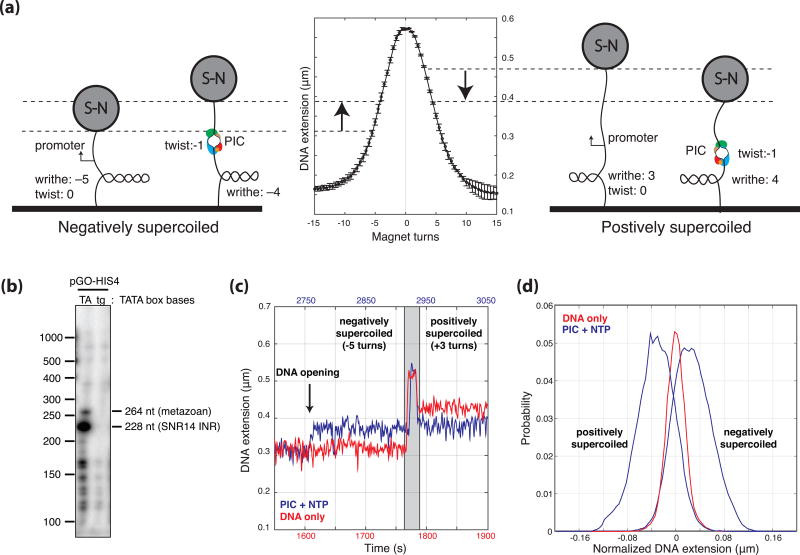
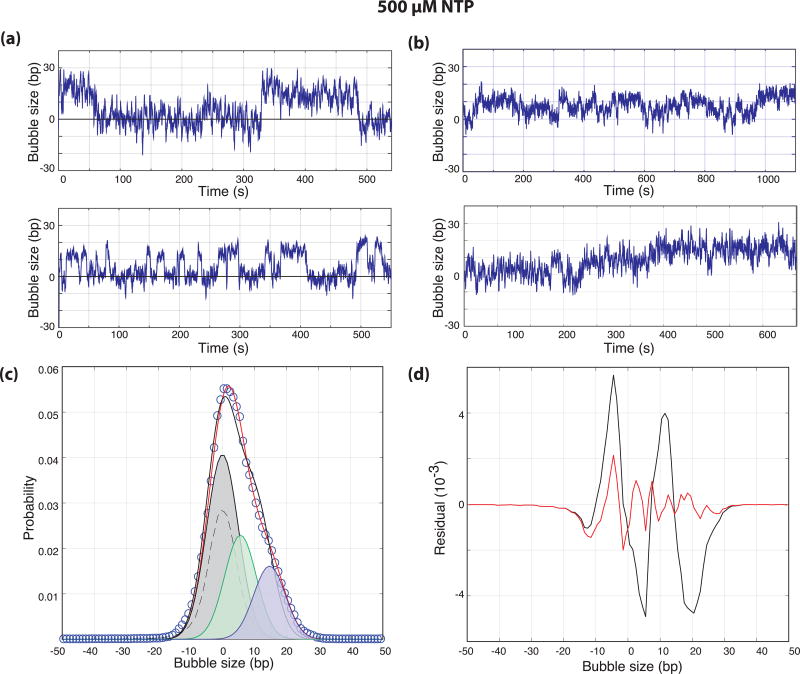
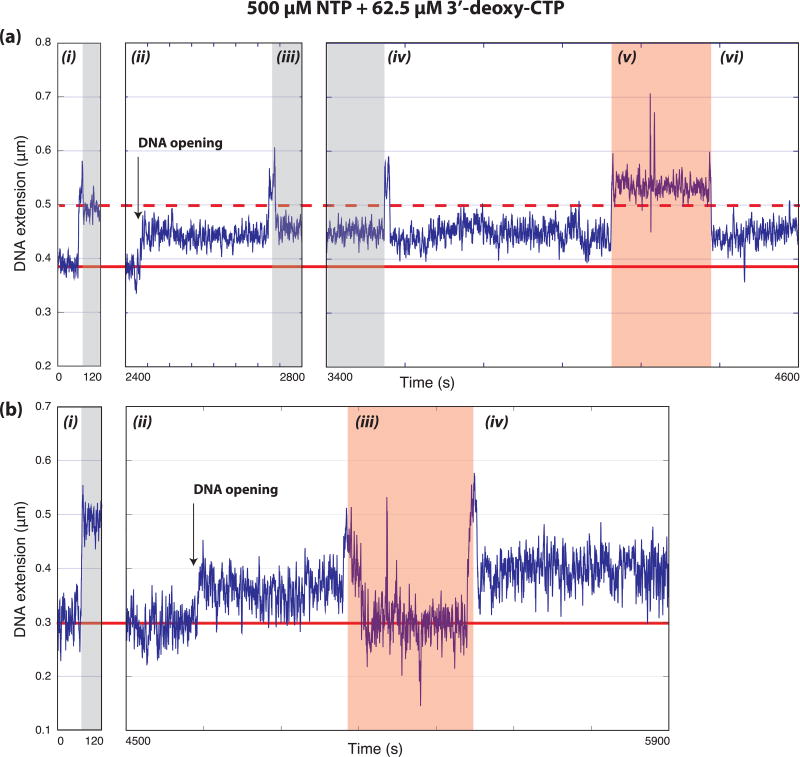
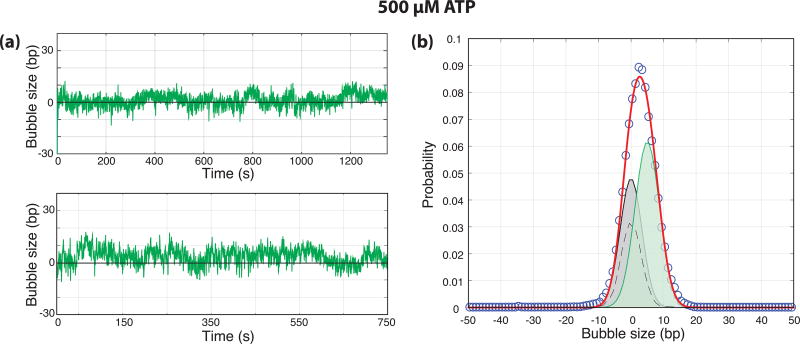
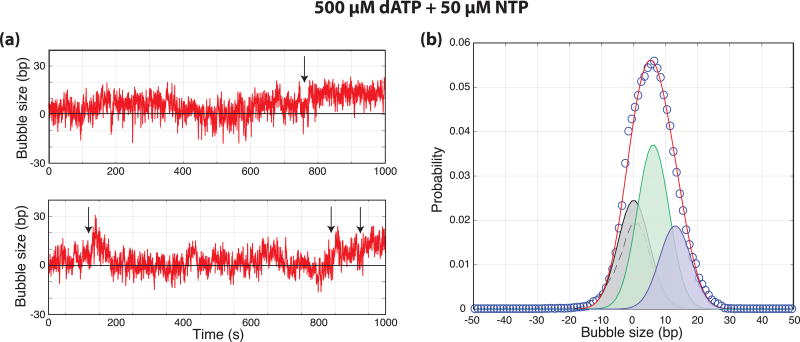
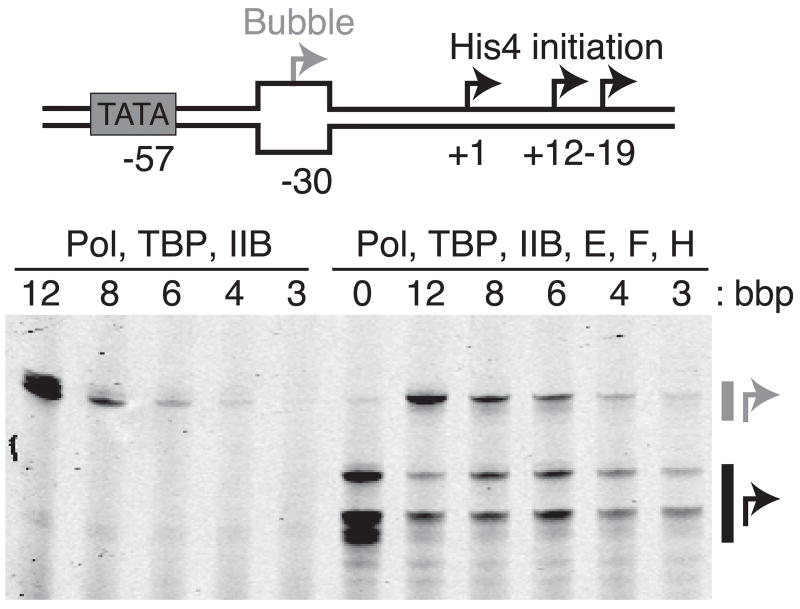
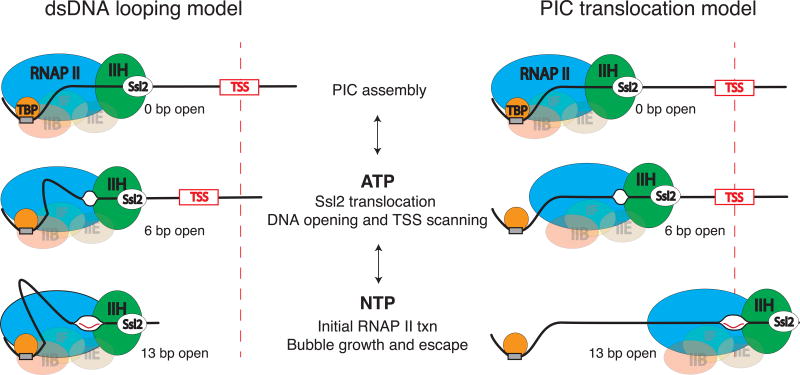
Eukaryotic mRNA transcription initiation is directed by the formation of the megadalton-sized preinitiation complex (PIC). After PIC formation, double-stranded DNA (dsDNA) is unwound to form a single-stranded DNA bubble, and the template strand is loaded into the polymerase active site. DNA opening is catalyzed by Ssl2 (XPB), the dsDNA translocase subunit of the basal transcription factor TFIIH. In yeast, transcription initiation proceeds through a scanning phase during which downstream DNA is searched for optimal start sites. Here, to test models for initial DNA opening and start-site scanning, we measure the DNA-bubble sizes generated by Saccharomyces cerevisiae PICs in real time using single-molecule magnetic tweezers. We show that ATP hydrolysis by Ssl2 opens a 6-base-pair (bp) bubble that grows to 13 bp in the presence of NTPs. These observations support a two-step model wherein ATP-dependent Ssl2 translocation leads to a 6-bp open complex that RNA polymerase II expands via NTP-dependent RNA transcription.
Figures
References
-
- Sainsbury S, Bernecky C, Cramer P. Structural basis of transcriptioninitiation by RNA polymerase II. Nat Rev Mol Cell Biol. 2015;16:129–143. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
